Биология - Учебно-практический справочник
Основы эволюционного учения - Историческое развитие органического мира
Проблема эволюции — центральная проблема биологии, однако сам подход к трактовке этого понятия отличается в разных теориях. Этимология слова «эволюция» идентична русскому «развитие», но используются эти слова в разных смыслах. Развитие чаще трактуют как онтогенез, изменения, происходящие поколение за поколением. Эволюция (историческое развитие) — изменение, которое приводит к чему-то новому. К характерным ее особенностям можно отнести возникновение новых форм, необратимость изменений, их прогрессивный характер или вырабатывание новых приспособлений.
Изучение феномена эволюции происходит с двух разных точек зрения. Науки об истории (палеонтология, филогенетика) изучают, как проходила эволюция. Механизмы этого процесса изучает другая наука, которую часто называют по-разному: эволюционная биология, эволюционное учение, эволюционная теория, эволюционизм и даже просто — дарвинизм.
Для периодизации истории эволюционной биологии мы используем принцип, предложенный известным российским эволюционистом Николаем Николаевичем Воронцовым (1934-2000). В истории эволюционного учения, согласно Воронцову, можно выделить три «синтеза».
Додарвиновский период был периодом до I синтеза. Его начало включало первые попытки объяснить целесообразность животных, сделанные еще в античности. К началу XIX в. биология накопила множество фактов, косвенно и прямо говоривших о феномене эволюции. Свидетельства об этих фактах надо было собрать воедино, рассмотрев в связи с единой теорией.
Гипотеза эволюции Жана Батиста Ламарка
Первая попытка обобщить объяснения изменений организмов во времени принадлежит французскому ученому Ж. Б. Ламарку (1744-1829), опубликовавшему в 1809 г. труд «Философия зоологии». Он утверждал, что все живые организмы под воздействием окружающей среды приобретают полезные изменения. Низшие организмы, не имеющие нервной системы, изменяются непосредственно под влиянием факторов внешней среды (листья водных растений удлиняются под влиянием течения воды и т. п.). Высшие организмы (в первую очередь животные) производят приспособления, тренируя определенные органы (вытягивание шеи жирафа, тянущегося за листьями). Органы, которые не тренируются, со временем исчезают.
Факторами эволюции Ламарк считал наследование каких-либо изменений организмом (в том числе и модификаций) и внутреннее стремление организмов к прогрессу. Сам эволюционный процесс ученый рассматривал как очередную смену — градацию, которая идет от низших стадий развития к высшим. При этом наличие низших форм жизни он объяснял непрерывным самозарождением. Современные представления, основанные на взглядах Ламарка, называются неоламаркизм.
Эволюционное учение Чарльза Дарвина — 1 синтез
Классический дарвинизм стал 1 синтезом в эволюционной биологии. Трудами Чарльза Дарвина и трактовками его идей учеными значительная часть биологии была перестроена в эволюционном ключе в течение 60-80 гг. XIX века.
Вряд ли кто-то повлиял больше на историю биологии, чем Чарльз Роберт Дарвин (1809-1882). В 1831-1836 гг. молодой Дарвин в качестве натуралиста принял участие в кругосветном путешествии на исследовательском судне «Бигль». Богатый материал, собранный им во время путешествия, знакомство с традицией трансформизма (в изложении его деда Эразма Дарвина), склонность к методическим размышлениям привели Дарвина к эволюционным идеям. В 1859 г. Дарвин представил на заседании научного общества свой доклад и статью Уоллеса, в которой тот независимо сформулировал подобные выводы. В том же году была опубликована известная книга «Происхождение видов путем естественного отбора, или Сохранение избранных рас в борьбе за жизнь». Изложенные в этой книге положения Чарльза Дарвина таковы. Единицей эволюции является отдельная особь. Особи одного вида отличаются друг от друга множеством признаков. Организмы размножаются в геометрической прогрессии, и любой из видов может быстро превысить свою предельную возможную численность.
Ограниченность ресурсов порождает борьбу за существование — выживание одних особей и гибель других. Дарвин выделял межвидовую и внутривидовую борьбу с факторами неживой природы. Эволюция происходит на основе наследственной изменчивости. Она не имеет приспособительного характера (неадаптивная). Механизм, обеспечивающий приспособление организмов к условиям окружающей среды, — естественный отбор, который заключается в преимущественном выживании и размножении более приспособленных особей. Этот термин Дарвин ввел как аналог искусственного отбора, используемый в селекции. Одной из форм естественного отбора является половой отбор, который описывает соперничество и преимущество одних особей перед другими в спаривании с особями другого пола.
Естественный отбор изолированных видов ведет к дивергенции (расхождению) их признаков и, в конце концов, приводит к видообразованию. Противоположным процессом является конвергенция. Даже идея естественного отбора высказывалась многими авторами задолго до Дарвина. Успех дарвинизма обусловило объединение разрозненных идей в единую теорию.
Основы эволюционного учения
Для объяснения наследования приобретенных признаков Дарвин публикует в 1868 г. (в книге «Изменения домашних животных и культурных растений») так называемую «временную гипотезу пангенезиса». Согласно этой гипотезе, все ткани организма должны направлять в половые клетки особые частицы — геммулы, передающие будущему поколению свойства, приобретенные отцом за время его жизни. В 1871 г. Дарвин публикует вторую по важности из числа своих произведений книгу «Происхождение человека и половой отбор». В ней, кроме детального обсуждения доказательств происхождения человека от общих предков с современными человекообразными обезьянами, выдвинута концепция полового отбора. Согласно этой концепции, если какие-то признаки самцов способствуют их привлекательности для самок, степень выраженности этих признаков в ряду поколений усиливается, даже если они снижают шансы каждой отдельной особи на выживание. Так, хвост самцов павлинов, хоть и делает их менее защищенными от хищников, должен эволюционировать благодаря тому, что самки охотнее спариваются с длиннохвостыми партнерами.
Впрочем, дарвинизм встретил сопротивление не только сторонников буквальной трактовки священных книг христианства, но и ряда серьезных специалистов, которые были сторонниками креационизма — представлений о неизменности видов и создании их Богом.
Дарвинизму сложно объяснить механизмы наследования благоприятных отклонений, возникновение принципиально новых признаков и эволюцию в определенном направлении. Эти и другие трудности привели к кризису эволюционной биологии.
Период кризиса 1 синтеза пришелся на конец XIX — начало ХХ века. Проявляется отсутствие представлений о механизмах наследственности и изменчивости. В это время происходит переоткрытие менделевских законов наследственности. Август Вейсман (1834-1914) доказал, что полученные в течение жизни особей изменения тела не наследуются, и назвал свою концепцию «неодарвинизмом». Вильгельм Людвиг Иогансен (1857-1927) показал, что без генетической изменчивости естественный отбор неэффективен. Появились разнообразные альтернативы классическому дарвинизму, в том числе сальтационизм (теория о скачкообразном характере эволюции) Гуго де Фриза (1848-1935), номогенез (теория о закономерном характере эволюции) Льва Семеновича Берга (1876-1950) и др.
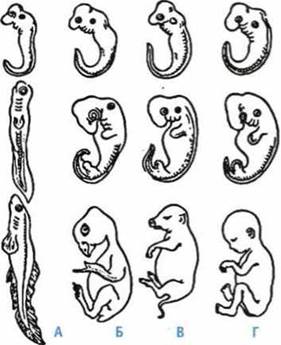
Сравнение стадий эмбрионального развития позвоночных животных:
А — рыба; Б —птица; В — свинья; Г — человек
Эрнстом Геккелем и Фридрихом Мюллером независимо друг от друга был открыт биогенетический закон, суть которого заключается в том, что онтогенез определенного организма повторяет филогенез данного вида. Этот закон наблюдается в сходстве строения эмбрионов позвоночных животных.
Филогенез — историческое развитие определенных систематических групп и вообще всего живого. Термин был предложен Геккелем, который для определения исторического развития определенной группы предложил метод тройного параллелизма: сопоставление данных палеонтологии, эмбриологии и сравнительной анатомии. Последовательность изменений организмов или их органов в пределах определенной систематической категории была названа филогенетическим рядом.
Благодаря работам английского ученого Т. Гексли, русского — А. О. Ковалевского, немецкого — К. Гегенбауэра и других были введены понятия, описывающие разницу и сходство в строении и функциях органов разных систематических категорий:
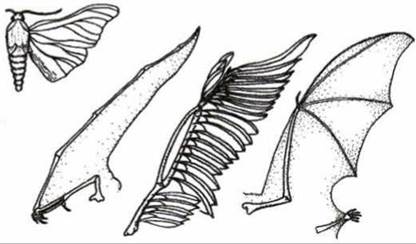
• аналогичные органы— органы, имеющие различное строение и происхождение, но выполняющие одинаковые функции (крыло птицы и бабочки);
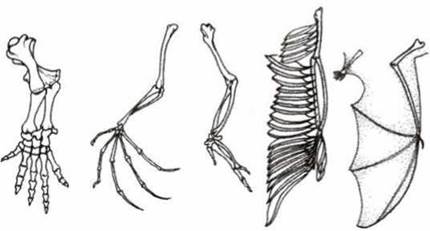
• гомологичные органы — органы, имеющие общее происхождение, но разные функции (рука человека, крыло летучей мыши, «плавник» кита);
• рудименты — органы с упрощенным строением по сравнению с формой предков вследствие потери в филогенезе своих функций (тазовый пояс китообразных, околоцветник злаков и т. п.);
• атавизмы — проявление у отдельных особей признаков, присущих форме предков (хвост у человека, недоразвитые конечности у веретеницы).
Аналогичные органы
Гомологичные органы
Определение приспособлений, их происхождение и функции имеют большое значение. В первую очередь ученых интересуют внешние характеристики: форма тела, окраска и т. д. Животные с защитной окраской и формой тела могут эффективно маскироваться под предметы окружающей среды (палочники). Яркая предупредительная окраска служит для информирования потенциальных хищников о наличии защитных механизмов (яд — божья коровка, жалящие структуры — осы). Демонстрационная или привлекательная окраска и поведение служат для отпугивания хищников или конкурентов, для привлечения опылителей или особей противоположного пола.
Особой категорией подобных адаптаций является мимикрия—способность к подражанию окраске и форме хорошо защищенных организмов плохо защищенными. Вид, которому подражают, называется моделью, а тот, который подражает, — имитатором. Выделяют две формы мимикрии:
• бейтсовская мимикрия (автор — английский энтомолог Г. Бейтс) заключается в том, что плохо защищенный вид подражает хорошо защищенному (мухи подражают осам);
• мюллеровская мимикрия (автор — немецкий зоолог Ф. Мюллер) заключается в формировании «кольца» защищенных видов, похожих друг на друга. При взаимодействии хищника с одним из видов он не будет трогать других (различные виды божьих коровок).
Синтетическая теория эволюции — II синтез
Начало II синтеза в эволюционной биологии, который получил название синтетической теории эволюции (СТЭ), было положено работой российского ученого Сергея Сергеевича Четверикова (1880-1959), который в 1926 г. показал, что учение о естественном отборе и генетика не противоречат одно другой. В работе утверждалось, что фенотипически нормальные особи из природных местообитаний являются носителями значительного генетического разнообразия. В серии работ ряда других авторов в дальнейшем была сформулирована концепция эволюции как следствия отбора генов. Важнейший вклад в эту концепцию внес Джулиан Хаксли (1887-1975), опубликовавший в 1942 г. монографию «Эволюция. Новый синтез».
Согласно СТЭ, эволюция — следствие комплексного процесса, рекомбинации генов и отбора носителей определенных генотипов. Единицей эволюции является популяция. Элементарное эволюционное событие, с этой точки зрения, — изменение аллельных частот в популяции. Именно это переводит популяцию в новое, по сравнению с ее историей, состояние. Создатели СТЭ предполагали, что мутации и рекомбинации носят произвольный характер, и только отбор придает эволюции определенное направление. Изменять соотношение аллелей в популяциях (по СТЭ) могут и случайные события — во-первых, дрейф генов и популяционные волны, во-вторых, эффект основателя (сокращение генетического разнообразия в популяциях, основанных небольшим количеством особей).
Важное условие для того, чтобы эволюция привела к принципиально новому событию — возникновению нового вида, — изоляция между популяциями.
Выделяют несколько ее форм:
• географическая — наличие между популяциями одного вида географических преград (река, иной тип биогеоценоза и т. п.);
• экологическая — формирование совокупности особей с различными требованиями к условиям окружающей среды (предпочтение определенного типа пищи, уровня влажности и т. п.);
• сезонная (временная) — наблюдается при размножении особей одного вида в разное время;
• этологическая (поведенческая) — зависит от особенностей поведения особей (различная песня самцов птиц);
• генетическая — несовместимость гамет, и, следовательно, невозможность оплодотворения или образования плодовитого потомства.
Факторами эволюции, согласно СТЭ, являются мутации, рекомбинации, отбор, дрейф генов и эффект основателя, поток генов, изоляция и естественный отбор.
Микроэволюция
Микроэволюция — эволюционные процессы, протекающие в популяциях определенного вида. Пластичность видов заключается в существовании подвидов, особи которых отличаются от других подобных совокупностей приспособлениями к конкретным условиям среды.
С точки зрения СТЭ, единственным источником наследственной изменчивости является мутагенез. Мутации — изменения генотипа, которые вызывают формирование фенов — элементарных наследственных изменений фенотипов как приспособлений к условиям окружающей среды, направляемых естественным отбором. В этом заключается творческая роль естественного отбора.
В зависимости от направления адаптивных изменений выделяют:
• стабилизирующий отбор — направлен на поддержание постоянства определенного фенотипа, сужает норму реакции. Проявляется в относительно постоянных условиях среды;
• движущий, или направленный отбор — способствует изменчивости определенного направления, сдвигая норму реакции в определенную сторону. Проявляется в условиях медленных изменений окружающей среды в определенном направлении или при расширении ареала;
• разрывающий, или дизруптивный отбор — направляет изменчивость в двух или более направлениях, подавляя проявление средних состояний признаков. Обеспечивает существование в нестабильных условиях окружающей среды (существование насекомых на океанических островах с постоянными ветрами).
Видообразование — эволюционный процесс образования новых видов. Имеет необратимый характер.
Выделяют географическое и экологическое видообразование. Видообразование является результатом соответствующей формы изоляции. Выделяют еще полиплоидизацию и скрещивание между особями близких видов без дивергенции (вид- потомок вытесняет родительский вид).
Макроэволюция
Макроэволюция включает эволюционные процессы, приводящие к возникновению таксонов надвидовых рангов (родов, семейств и т. д.). Считается, что виды одного рода, роды одного семейства и т. д. имеют единого общего предка (принцип монофилии). Разнообразие же видов, которое возникает при приспособлении к условиям среды, называется адаптивной радиацией.
Изучая историческое развитие преимущественно позвоночных, Н. Северцов предложил гипотезы биологического прогресса и регресса. Биологический прогресс сопровождается увеличением численности популяций и расширением ареала. Он является результатом эволюционного успеха определенного вида. Биологический perpecc характеризуется уменьшением численности популяций и сужением ареала. Он является следствием неспособности приспособиться к условиям среды. Может привести к вымиранию определенной группы организмов.
Существует несколько эволюционных механизмов — путей достижения биологического прогресса. Ароморфоз приводит к повышению уровня организации в целом и дает возможность вырабатывать новые приспособления к условиям среды (возникновение челюстей у рыб, преобразования в скелете птиц для полета и т. д.).
Дегенерация (морфофизиологический регресс) — упрощение строения организмов в процессе эволюции (эндопаразиты, малоподвижные организмы — покровные, морские желуди и др.). Идиоадаптацкя — преобразования организмов, которые не изменяют уровень его организации (разнообразие окраски, строения цветков и т. п.).
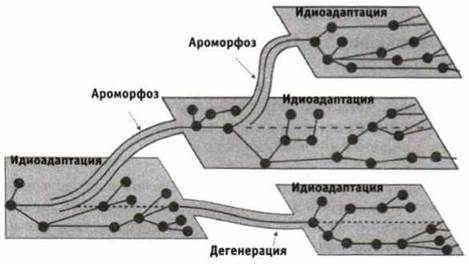
Схема соотношения между ароморфоэом, идиоадаптацией и дегенерацией
Дальнейшее развитие теории эволюции
Кризис II синтеза, связанный с накоплением противоречивых фактов, начался еще в эпоху формирования теории. Например, еще в 1930-е годы Джон Б. С. Холдейн (1892-1964), один из создателей СТЭ, показал, что естественный отбор по многим парам аллелей одновременно оказывается совершенно неэффективным.
Критика СТЭ достигла значительной остроты в конце ХХ века. Развитие биологии привело к существенному пересмотру практически всех положений этой теории. В противовес СТЭ развивается целый ряд альтернативных теорий эволюции. В числе важнейших следует назвать неосальтационизм Рихарда Гольдшмидта (1878-1958), экосистемную теорию эволюции Валентина Абрамовича Красилова, теорию нейтральной эволюции Мото Кимуры (1924-1994), теорию прерывистого равновесия Стивена Гулда (1941-2002) и других американских палеонтологов, эпигенетическую теорию эволюции (ЭТЭ) Михаила Александровича Шишкина и др.
Совершенствование теории эволюции далеко не закончено. За последние два десятилетия значительно возросло количество отечественных и зарубежных публикаций, в которых отмечается, что синтетическая теория не до конца адекватна современным знаниям о ходе эволюционного процесса.
Наряду с закономерностями, описываемыми СТЭ, сохраняются проблемы, требующие объяснения. К ним относится, например, проблема ускорения выработки морфологических приспособлений с ходом эволюции. В их числе — быстрые эволюционные изменения, вызванные гибридизацией и «горизонтальным» переносом генетической информации (т. е. передачей информации между неродственными группами организмов), ролью симбиоза и эндосимбиоза в эволюции.
Другой пример связан с концепцией «прерывистого равновесия». Она основана на простом палеонтологическом наблюдении: продолжительность периодов видового постоянства на несколько порядков превышает длительность переходов из одного состояния в другое. Судя по имеющимся данным, это правило в общем справедливо для всей ископаемой истории многоклеточных животных и имеет достаточное количество подтверждений.
Как одно из наиболее часто критикуемых общих положений синтетической теории эволюции можно привести ее подход к объяснению вторичного сходства, т. е. одинаковых морфологических и функциональных признаков, которые не были унаследованы, а возникли независимо в разных линиях эволюции организмов.
Еще недостаточно осмыслены эксперименты, которые показали, что отбор в направлении определенных фенотипических реакций и изменения условий среды могут делать эти реакции устойчивыми и развивающимися без специфических внешних воздействий. Открыт ряд новых молекулярно-биологических механизмов, которые показывают возможность направленной перестройки со стороны организма генетической информации (роли малых РНК, парамутаций, мобильных генетических элементов и т. п.). Вероятно, интеграция новых данных в новой теории — дело ближайшего будущего. Возможно, конец ХХ — начало XXI в. впоследствии назовут периодом, когда формировался III синтез в эволюционной теории.