Биогеография общая и частная: суши, моря и континентальных водоёмов - 2017 год
Озеро Байкал - Специфика биот крупных озёр - Общая характеристика континентальных водоёмов как биотопа - Биогеография континентальных водоёмов
Условия жизни в крупных озёрах обычно весьма сильно отличаются от условий жизни в мелких. Площади крупных озёр превышают площади речных ЦС (естественно, самих водотоков, а не водосборных бассейнов), а постоянство условий принципиально выше. Время существования большинства озёр — всего лишь несколько тысячелетий, крупные же озёра существуют часто миллионы лет. Монографии, опубликованные по двум крупным озёрам — Байкалу (Кожов, 1962; Кожова, Изместьева, 1998) и Танганьике (Coulter, 1991), — позволяют рассмотреть на их примере особенности биот крупных озёр. Приведённые ниже характеристики основаны на этих обзорах (ссылки на них часто опущены) и дополнены данными, взятыми из отдельных статей и более частных обзоров.
Озеро Байкал
В Байкале содержится около 20% всей жидкой пресной воды Земли (столько же, сколько в водоёмах и водотоках Амазонки). Возраст Байкала оценивают в 28—24 млн. лет. Это — одно из древнейших озёр Земли, больший возраст имеют только Эри (20—50 млн. лет) и Маракайбо (36 млн. лет) (Roth, 2009). Возникновение Байкальского рифта, вероятно, синхронно возникновению рифтовой системы восточной Африки, где ныне располагаются озёра Танганьика, Ньяса и ряд более мелких. Озёра на территории современного Байкала и его окрестностей существовали постоянно, но это были мелкие озёра, биота которых не отличалась от биот окрестных водоёмов. Глубоководные озёра глубиной в несколько сотен метров начали возникать в начале олигоцена (не ранее 30 млн. лет назад). Они пережили сложную историю объединений и изоляций, сопровождавшихся значительными колебаниями их уровня. Палеонтологические данные показывают, что в плиоцене, а, возможно, даже в миоцене байкальская фауна была экологически и таксономически отлична от фауны мелководных сибирских озёр. Это означает, что Байкал уже существовал как большое и глубокое озеро (или система таких озёр). На границе плиоцена-плейстоцена (2—1,5 млн. лет назад) прогрессирующее похолодание климата привело к появлению льда зимой, т. е. аэрации всей толщи воды озера до дна, сделав возможной возникновение глубоководной биоты. В плейстоцене менялась протяжённость ледового периода, но постоянного ледового покрова не было.
В настоящее время Байкал — озеро бассейна Енисея, но присоединение Байкала к этому бассейну произошло недавно, в раннем плейстоцене (800 тыс. лет назад), до этого он принадлежал бассейну Лены. Байкал летом прогревается до 12—16°С, зимой замерзает, поэтому вертикальное перемешивание охватывает всю его толщу и высокая концентрация кислорода (до 75%) сохраняется до самого дна на протяжении всего года. Впрочем, деятельность Байкальского целлюлозно-бумажного комбината привела к образованию обширных площадей, покрытых лигнином и лишённых кислорода. Летом термоклин в открытой части находится на глубине, по крайней мере, 10—20 м, зимой — занимает толщу воды 50—100 м. Сезонные колебания температуры прослеживаются до глубины 300 м. В открытых частях озера придонная температура редко превышает 12—13°С и то лишь на несколько дней в августе. В сорах и мелких закрытых и полузакрытых заливах придонная температура достигает 22—23° C и превышает характерные для озера летом 12—13°C, по крайней мере, 3,5 месяца в году.
Фауна озера образована более чем 2500 видами, преимущественно эндемичными. Но эта цифра характеризует биоту озера в целом. Анализ ареалов видов показывает, что в географических пределах озеро населено двумя разными биотами. Мелководные хорошо прогреваемые заливы (соры) и дельты впадающих в Байкал рек, географически относимые к озеру, биогеографически и экологически к нему не принадлежат. Они населёны ценозами, не отличающимися от ценозов окрестных сибирских озер, существующие в условиях идентичных тем, к которым они адаптированы. Эндемичные виды тут практически отсутствуют.
Остальная (большая) часть Байкала населена эндемичными ценозами. В их составе многие обычные и разнообразные в окрестных водоёмах группы редки, малоразнообразны или вовсе отсутствуют: подёнки, веснянки, стрекозы, жуки, клопы-водомерки (заменены на эндемичных бескрылых ручейников Рис. 4.3) и скорпионы, Chironomidae, наяды****** (единственный встречающийся в Байкале вид — Colletopterum ponderosum обитает только в сорах (Слугина, Старобогатов, 1999), дышащие лёгкими гастроподы Viviparidae, Decapoda, Branchiopoda.
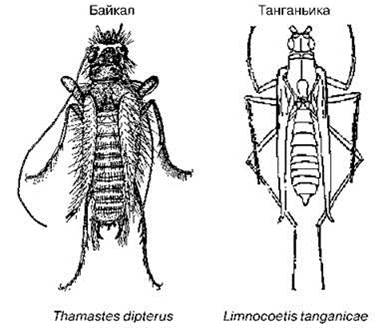
Рис. 4.3. Пелагические виды ручейников озер Байкал и Танганьика, связанные с поверхностной плёнкой. По: Кожов (1962); Heckey (1991), Tobias, Tobias (2008) и Marlier (1955).
Основу эндемичных ценозов создают Gammaroidea, донные Cyclopoida, Harpacticoida, Ostracoda, Nematoda, Oligochaeta, Bathynellidae, бычки Cottoidei и многие другие таксоны, для них характерно наличие большого числа эндемов. Разнообразны в Байкале бокоплавы, 363 эндемичных вида и подвида которых описаны из самого озера и ещё 12 из вытекающих из него рек, это в полтора раза больше, чем известно видов бокоплавов из Южной Америки, Африки, тропической Азии и Австралии вместе взятых (Рис. 4.37). Эти виды относятся к 72 родам и 7 эндемичным семействам (Камалтинов, 2002), образовавшимся, вероятно, из голарктического Gammarus (Englisch et al., 2003; Macdonald et al., 2005). Ихтиофауна озера включает 56 видов и подвидов рыб 14 семейств (Sideleva, 1994), из них 37 эндемичных видов (Павлов, Остроумов, 1998). Примечателен букет видов бычков Cottotoidei, состоящий из 29 эндемичных видов (11 родов). Анализ их возраста методом генетических часов показывает, что они начали дивергировать только в плейстоцене, не ранее 2 млн.л.н. (Sherbakov, 1999). Высокий эндемизм свойствен и другим группам. В то же время насекомых, доминирующих по числу видов в окрестных водоёмах, кроме 17 видов эндемичных ручейников в Байкале нет.
Тщательное изучение распределения отдельных видов показывает, что, хотя некоторые активно передвигающиеся сибирские виды могут быть найдены в байкальском биогеографическом районе, а наиболее эврибионтные байкальские эндемы выходят за его пределы, граница биогеографического региона, населённого эндемичной байкальской биотой, весьма резка.
Предложено несколько объяснений существования столь резкой границы между байкальской и сибирской биотами:
— изменение строения воды из-за большой глубины озера (Верещагин, 1935, 1940b; Бирштейн, 1946; Берг, 1949б);
— отличия в температуре воды и содержании в ней кислорода;
— взаимодействие голарктической и байкальской фаун (Леванидова, 1948).
Первое объяснение ныне можно воспринимать лишь как курьёз. Относительно второго можно отметить, что действительно, многочисленные эксперименты по выживаемости и размножению видов обоих биот (обзор см. Кожова, Изместьева, 1998) показывают, что для байкальских видов, в отличие от сибирских, предпочтительна более низкая температура и более высокое содержание кислорода. Как обычно, в таких экспериментах была также показана разная степень стенобионтности отдельных видов. Поэтому различие в аутоэкологии отдельных видов не способно объяснить, почему граница между комплексами видов столь резка. Лишь третье объяснение объясняет все факты. Биогеографическая граница проходит не по глубине, придонной температуре и т. п., а по градиенту комплекса абиотических факторов и разделяет акватории, в которых та или иная ЦС получают конкурентные преимущества, поскольку их экологические амплитуды различны. Поэтому анализ распределения видов по одному из факторов (например, по глубине) даёт, как обычно, искажённую картину, размывая эту биогеографическую границу. Более того, как было показано выше, для существования такой границы вовсе не обязательны различия ни в аутоэкологии, ни в абиотических факторах.
В Главе 1 было сказано, что многие исследователи объясняют высокий эндемизм байкальской биоты гипотезой реликтов, суть которой в том, что Байкал — рефугиум ранее широко распространённой биоты. Но существует и противоположное мнение, согласно которому эндемичная байкальская биота имеет местное происхождение (Андрусов 1903; Дорогостайский, 1923; Ламакин, 1955). Очевидно, что аллопатрическое видообразование не было главным способом видообразования, хотя и могло иметь место, поскольку ранее в районе Байкала было несколько крупных глубоководных озёр. О.М. Кожова и Л.Р. Изместьева (1998: 190—193) приводят многочисленные примеры несомненного происхождения глубоководных и пелагических видов от мелководных бентических. По их мнению в этом случае аллопатрический путь видообразования совершенно невозможен.
Анализ видовых ареалов эндемичной биоты показывает, что в Байкале можно выделить три биогеографических района: пелагический и два бентических: мелководный (в эпилимнионе) и глубоководный (в гиполимнионе).
Для озёр, прежде всего, специфично наличие пелагиали. Видообразование среди рыб начинается с образования эндемичных пелагических рас, которые в зависимости от особенностей наличных ресурсов могут превратиться в новые виды, рода или семейства, вероятно, сходно это происходит и в других таксонах. Пелагиаль Байкала в полной мере соответствует этой закономерности (Рис. 4.4). Основу первичной продукции Байкала дают диатомовые, из которых наибольшую роль играет эндемичная Aulacoseirabaicalensis (ранее её относили к роду Melosira); роль её столь велика, что говорят о “мелозирных” годах. Первая вспышка фитопланктона происходит до вскрытия льда, когда ещё сохраняется обратная стратификация, а количество света резко возрастает. В этот период создаётся порядка 80% органического вещества (Экосистема..., 1993). После разрушения ледового покрова разрушается и стратификация, что не позволяет возникать высоким концентрациям фитопланктона. Следующие вспышки (менее интенсивные) происходят после возникновения прямой стратификации, их обеспечивают синезелёные (летом) или диатомовые (осенью) водоросли. Суммарная первичная продукция фитопланктона Байкала составляет 127 гС/кв. м в год (Растительный мир..., 1978), т. е. Байкал является мезотрофным или даже эвтрофным озером (см. Таблицу 4.6). В зоопланктоне отчётливо доминирует растительноядный эндемичный веслоногий рак эпишура Epischura baicalensis (Calanoida), дающий до 90% биомассы и численности зоопланктона и эврифаг бокоплав Macrohectopus branizkii (Тереза и др., 2006). M. branizkii принадлежит к эндемичным монотипическим Macrohectopidae. Внешне он напоминает мизиду или эуфаузииду (Рис. 4.4) и обитает исключительно в пелагиали, причём среди 363 эндемичных видов и подвидов бокоплавов Байкала это — единственный пелагический вид. Следующий трофический уровень образован рыбами, основной поток энергии идёт через исключительно пелагических голомянок Comephoridae и более прибрежных, но тоже преимущественно пелагических бычков-желтокрылок Cottocomephorus, которых, хотя и относят к бычкам Cottidae, но выделяют в отдельное подсемейство Cottocomephorinae, к которым относят ещё два бентических рода. Голомянки (два вида: большая Comephorus baicalensis и малая C. dybowskii) (Рис. 4.4), несомненно, также являются специализированной ветвью Cottidae. Малая — планктофаг, большая — хищник. Оба вида голомянок предпочитают гиполимнион, это — единственные рыбы, обычные там, остальные предпочитают эпилимнион, лишь омуль изредка спускается до глубины 200 м. Голомянки предпочитают эпилимнион лишь на ранних стадиях развития. Кроме морфологических особенностей, их отличает от бычков и способ размножения. Если бычки откладывают икру на дно, то голомянки перешли к живорождению. Бычков- желтокрылок тоже два вида и они принадлежат к роду, обитающему только в пелагиали Байкала. Венчает трофическую сеть Байкала (Рис. 4.5) омуль Coregonus autumnalis migratorius и эндемичный вид нерпы Pusa sibirica. Таким образом, пелагическая биота, хотя и немногочисленна, но её эндемы имеют наиболее высокий таксономический ранг, вплоть до семейства (голомянки Comephoridae, бокоплавы Macrohecto- pidae). Ни для одной из остальных ЦС Байкала не свойствен эндемизм столь высокого ранга, вероятно, переход к жизни в пелагиали требует существенных перестроек морфологии и биологии. Ведущие таксоны пелагиали голарктических озёр: коловратки, веслоногие раки Cyclopoida, Cladocera — не играют сколь-нибудь существенной роли в открытой пелагиали Байкала: из 32 эндемичных видов коловраток только один (Synchaeta pachypoda) обычен там, а среди 23 эндемичных видов Cyclopoida и 8 Cladocera таких вовсе нет. Cladocera встречаются только в некоторых заливах и летом в прибрежных районах (Афанасьева, Шамараев, 2006). Примечательно, что карповые Cyprinidae и окуни Percidae, которые часто образуют пелагические таксоны, в Байкале не входят в состав пелагической ЦС. В целом пелагическая ЦС Байкала включает весьма небольшое число видов, многие из них имеют близких видов в больших реках и горных озёрах Евразии (Rotifera, Cyclops, Epischura).
Табл. 4.6. Вертикальная зональность Байкала по разным авторам.
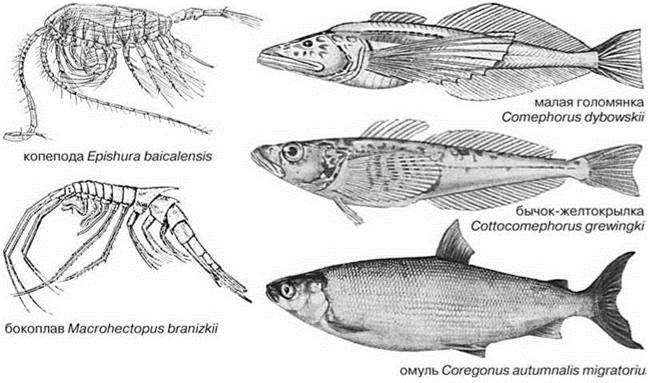
Рис. 4.4. Основные виды животных пелагиали Байкала (из: Кожов, 1962).
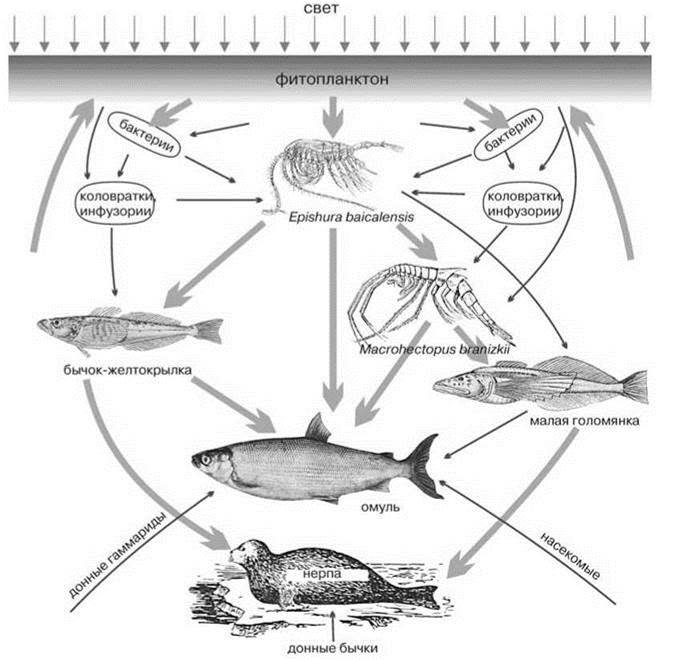
Рис. 4.5. Схема пищевых связей пелагиали Байкала (по Кожов, 1962).
Толстые линии — главные связи, тонкие — второстепенные.
В бентали разные авторы выделяли разное число вертикальных зон (районов), обзор дан М.М. Кожовым (1962) (Табл. 4.7). Выделение этих зон, однако, проводят на основе гидрологии, геоморфологии, состава грунтов и многим другим параметрам, но только не по составу биоты, в лучшем случае нам лишь сообщают, какие виды характерны для тех или иных зон, причём зачастую одни и те же виды указывают как характерные для разных зон. М.М. Кожов (1962) постоянно указывает на плохую изученность распределения биоты при характеристике каждой выделенной им зоны: сублитораль — “распределение животных ... изучено недостаточно”, супраабиссаль — “донное население. изучено слабо”, абиссаль — “бентос. исследован... ещё очень недостаточно” (стр. 172); а при характеристике нижней подзоны абиссали он пишет: “фауна. состоит почти сплошь из своеобразных глубоководных форм, большая часть из которых перечислена выше” (стр. 173), т. е. в характеристиках вышележащих зон. Даже авторы последней сводки по Байкалу (Кожова, Изместьева, 1998) выделяют зоны преимущественно по тем же гидрологическим критериям, лишь за границу сублиторали—супраабиссали принимают предел распространения бентосных диатомовых. Конечно, уклон дна озера, достигающий 40—60° и даже более, и обилие площадей, занятых выходами скальных пород, очень затрудняют изучение бентоса. Фактически ни одно из стандартных орудий лова бентоса в таких условиях не работает, исследования же с помощью подводных аппаратов (“Миров”) эпизодически и преследуют совершенно иные цели. К тому же дночерпатели очень плохо улавливают весьма разнообразных в озере бокоплавов и рыб, которые имеют весьма неравномерное распределение. Накопленные знания особенностей разных глубин не позволяют составить биологически обоснованную схему вертикальной зональности озера. На данном уровне наших знаний по составу биоты можно выделить, по-видимому, не более двух вертикальных зон: мелководную и глубоководную. Мелководная ЦС населяет эпилимнион. Большая часть эндемов озера входит в её состав. Глубоководная ЦС населяет глубины гиполимниона и сравнительно малоразнообразна и состоит исключительно из эндемов. Глубоководная зона, по-видимому, гомогенна, а мелководную, возможно, следует разделить на две*******: северную и южную (Кожова, Изместьева, 1998), но различия эти незначительны, а, возможно, и несущественны.
Итоговая схема биогеографического районирования Байкала следующая: (1) прибрежный район (мелководные заливы, соры и т. п.), (2) пелагиаль открытого озера, (3) мелководный бентический район (возможно, его следует поделить на северный и южный) и (4) глубоководный бентический район. Прибрежный район является частью окружающего озеро биогеографического района, тогда как остальные следует рассматривать как биогеографический регион высокого ранга (вероятно, подобласти). При этом все бентические районы следует противопоставить пелагическому, что можно отразить, присвоив бентали и пелагиали ранги провинций.