Биогеография общая и частная: суши, моря и континентальных водоёмов - 2017 год
Голарктика/Палеарктика/Неоарктика - Статус и границы некоторых территорий - Схемы биогеографического районирования суши - Биогеография суши
Суша, в отличие от океана, разделена на изолированные материки и острова. Распад суши произошёл в мезозое (подробно геологическая история суши в кайнозое рассмотрена в Главе 4). Фрагментация суши привела к тому, что биоты отдельных массивов суши в тропиках и умеренных широтах Южного полушария, и, даже до некоторой степени Северного, формировались в значительной степени независимо. Изоляция отдельных массивов суши определила особенности биот материков и островов. На территории каждого из них происходило образование СС из имеющегося набора таксонов. В разных регионах разные таксоны дали адаптивную радиацию для потребления сходных ресурсов. В кайнозое сухопутные связи возникли вновь, что позволило мигрировать представителям многих таксонов с одного массива суши на другой. В результате резко возросло сходство биот разных частей Голарктики, существенно уменьшились отличия Азии и Африки.
Предложено довольно много биогеографических схем суши*. В зоогеографии наибольшее распространение получили схемы, основанные на сходстве состава фаун. Большая часть из них восходит к схеме для птиц из шести областей, предложенной PL. Sclater в научном докладе, прочитанном перед Линнеевским обществом в 1857 г. и опубликованная в 1858 г. В схеме P.L. Sclater территории одного ранга имеют примерно одинаковое число эндемичных семейств. Схема была модифицирована Альфредом Уоллесом в 1876 г. для позвоночных в целом (Рис. 2.29). Невзирая на то, что Альфред Уоллес признавал несовершенство и ограниченность этой схемы, почти все остальные схемы так или иначе являются её преемниками (ср. Рис. 2.29 с Рис. 3.26). Это связано с тем, что P.L. Sclater и Альфред Уоллес, располагая ограниченными данными, верно подметили общие тенденции. Кроме этих схем были предложены и другие схемы, порой существенно отличающиеся. Как показано выше в Главе 2, схемы районирования в первую очередь определяют взгляды их авторов на то, как следует интерпретировать одни и те же фактические данные, что эти схемы должны отражать в первую очередь. Во вторую очередь на результаты влияют методы выделения регионов. В третью — методы их группировки и построения иерархии.
Использование более сложного математического аппарата само по себе не делает схемы более предпочтительными и даже адекватными. Например, схема Smith (Рис. 3.29), как и многие другие схемы построена в результате анализа современного распространения семейств млекопитающих, но с использованием многомерного шкалирования. Но мало кто предпочтёт её схемам, основанных на тех же данных (Рис. 3.26—3.28), где использование математики было ограничено арифметическими действиями. Если предлагаемые биогеографические схемы построены на основе данных по одному или нескольким таксонам, то особенности этих таксонов также может повлиять на результат. Схемы на Рис. 3.26—3.28 построены на основе данных по позвоночным (преимущественно птицам и млекопитающим), а схема Рис. Ц5 — по насекомым.
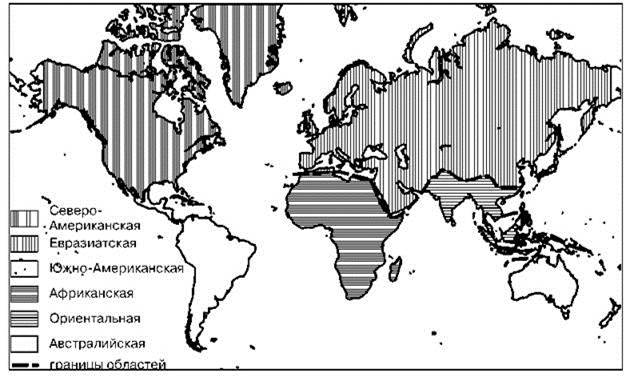
Рис. 3.26. Фаунистические области суши Cox (2001).
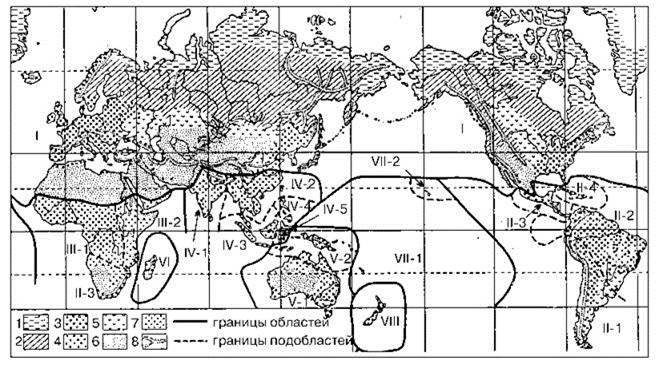
Рис. 3.27. Схема фаунистического районирования суши Бобринского (1946, 1951).
Области (подобласти): I — Голарктическая; II — Неотропическая (1 — Патагонско-Чилийская, 2 — Бразильская, 3 — Центральноамериканская, 4 — Антильская); III — Эфиопская (1 — Западноафриканская, 2 — Восточноафриканская, 3 — Южноафриканская), IV — Индо-Малайская (1 — Индийская, 2 — Бирманско-Китайская, 3 — Зондская, 4 — Филиппинская, 5 — Целебесская); V — Австралийская (1 — Австралийская, 2 — Папуасская); VI — Мадагаскарская; VII — Полинезийская (1 — Полинезийская, 2 — Гавайская); VIII — Новозеландская.
Ландшафты: 1 — тундра, 2 — тайга, 3 — леса широколиственные и субтропические, 4 — влаголюбивый тропический лес, 5 — степь, 6 — пустыня, 7 — саванна, 8 — горы.
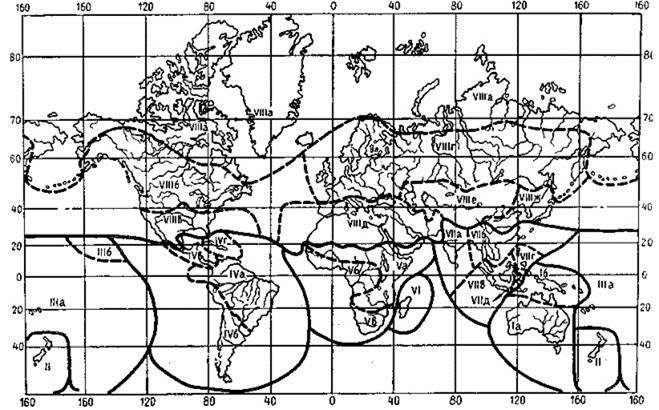
Рис. 3.28. Зоогеографические области (и подобласти) суши по Петрову (1966).
I — Голарктическая область (а — Арктическая, б — Канадская, в — Сонорская, г — Европейско-Сибирская, д — Средиземноморская, е — Центральноазиатская, ж — Маньчжурско-Китайская); II — Неотропическая (а — Гвиано-Бразильская, б — Чилийско-Патагонская, в — Центральноамериканская, г — Антильская); III — Эфиопская область (а — Восточноафриканская, б — Западноафриканская, г — Южноафриканская); IV — Восточная, или Индо-Малайской область (а — Индийская, б — Индо-Китайская, в — Зондская, г — Филиппинская, д — Целебесская); V — Австралийская область (а — Новоголландская, б — Папуасская (Новогвинейская); VI — Мадагаскарская область; VII — Новозеландская область; VIII — Полинезийская область (а — Собственно полинезийская, б — Гавайская). Материк Антарктики и ряд островов Южного Ледовитого океана лишены позвоночных животных, не связанных с морем, в связи с чем эти территории по включены в схему зоогеографических подразделений суши. Жирные линии — границы областей, пунктирные — подобластей.
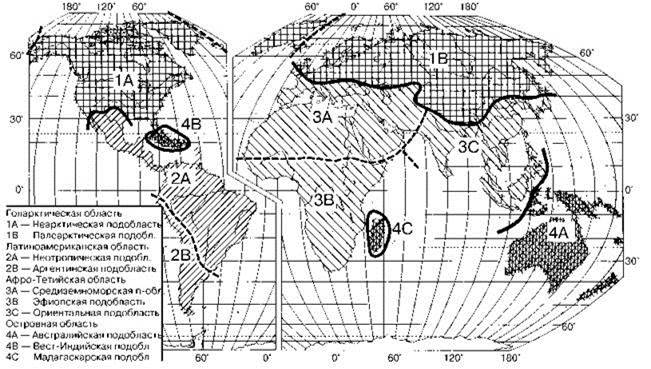
Рис. 3.29. Схема фаунистического районирования суши Smith (1983).
Фитогеографические схемы имеют определённые отличия от зоогеографических. В большинстве фитогеографических схем принято выделять шесть наиболее крупных областей, но границы их частью совпадают, частью очень сильно отличаются от принятых в зоогеографии. Традиции выделения этих царств восходят к середине XIX в. Сравнительно недавно эту схему детализировал А.Л. Тахтаджян (Рис. 3.30), предложивший схему фитогеографического районирования, описанию которой посвящены две его монографии, 1978 г. на русском и 1986 г. на английском, что обусловило её широкую известность не только у нас (Григорьева, 2014), но и за границей. Выше, при рассмотрении методов биогеографии (Глава 2) уже было показано, что теоретические представления, положенные, по утверждению автора, в её основу, сам он при её построении не использовал. Возможно, прав С.М. Разумовский, писавший, что “А.Л. Тахтаджян первоначально произвольно выделил свои фитохории, основываясь на традиции и интуитивных представлениях, а затем, как мог, сообщил число эндемичных таксонов, попавших на территории этих субъективных единиц” (1999: 31). Например, в схеме А.Л. Тахтаджяна те же шесть наиболее крупных подразделений, что и использовали в фитогеографии и до него (ср. со схемой Good (1973) на Рис. 3.32). Критерии, на которых основан ранг выделов разные для разных выделов. Так, например, для Судано-Замбезийской провинции характерно “три... эндемичных семейства... и сравнительно немного эндемичных родов, но число эндемичных видов велико” (Тахтаджян, 1978: 145). Эту провинцию он разделяет на четыре подпровинции, из которых по его же данным для трёх характерно небольшое число эндемичных родов, при полном отсутствии эндемичных семейств, зато в подпровинции о. Сокотра имеется эндемичное семейство и около 25 эндемичных родов. Сам С.М. Разумовский (Рис. 3.34), напротив, подробно объяснил, как им были получены предлагаемые выделы ранга района, округа и провинции. Объединять провинции в регионы более высокого ранга он предлагал на основе их “древних флорогенетических связей”. Каждая из флорогенетических областей соответствует материку Лавразийская область — Лавразии, а Гондванское — Гондване за вычетом Антарктиды, Антарктическое царство занимает молодую сушу и, может быть, занимало юг Гондваны (Антарктиду). Очевидно, что границы областей совершенно не обусловлены климатом — он одинаков по обе стороны границы по всем мыслимым показателям. Схемы флористического районирования на Рис. 3.30—3.32 широко используют в англоязычной литературе. Недавно схема Тахтаджяна была модифицирована Камелиным (2012) (Рис. 3.33).
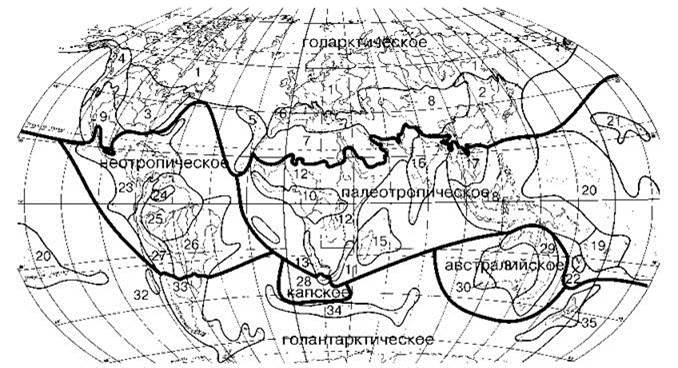
Рис. 3.30. Схема флористического районирования суши Тахтаджяна (1986, мало отличающиеся от его схем 1974 и 1978 гг.).
Названия областей надписаны, провинции: 1 — Циркумбореальная, 2 — Восточная азиатская, 3 — Североамериканская атлантическая, 4 — Скалистых гор, 5 — Макаронезийская, 6 — Средиземноморская, 7 — Сахаро-Аравийская, 8 — Ирано-Туранская, 9 — Мадреанская, 10 — Гвинео-Конголезская, 11 — Узумбаро-Зулулэндская, 12 — Судано-Замбезийская, 13 — Кару-Намиба, 14 — островов Св. Елены и Вознесения, 15 — Мадагаскарская, 16 — Индийская, 17 — Индокитайская, 18 — Малезийская, 19 — Фиджийская, 20 — Полинезийская, 21 — Гавайская, 22 — Новокаледонская, 23 — Карибская, 24 — Гвианского нагорья, 25 — Амазонская, 26 — Бразильская, 27 — Андийская, 28 — Капская, 29 — Северо-восточноавстралийская, 30 — Юго-западноавстралийская, 31 — Центральноавстралийская, 32 — Хуан-Фернандесская, 33 — Чилийско-Патагонская, 34 — Субантрактических островов, 35 — Новозелендская.
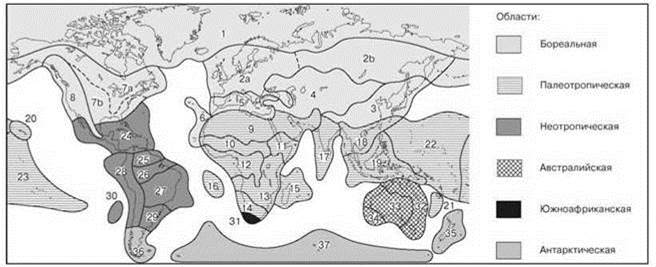
Рис. 3.31. Схема флористического районирования суши Good (1973).
Провинции: 1 — Арктическая и субарктическая, 2 — Евро-Сибирская (а — Европейская, b — Азиатская), 3 — Сино-Японская, 4 — Западная и Центральноазиатская, 5 — Средиземноморская, 6 — Макаронезийская, 7 — Атлантическая североамериканская (а — Северная, b — Южная), 8 — Тихоокеанская североамериканская; 9 — Африкано-индийская пустынная, 10 — Суданская парковая степная, 11 — Северо-восточная африканская высокогорная, 12 — Западноафриканская дождевого леса, 13 — Восточноафриканская степная, 14 — Южноафриканская, 15 — Мадагаскарская, 16 — островов Вознесения и Св. Елены, 17 — Индийская, 18 — Континентальная юго-восточноазиатская, 19 — Малазийская, 20 — Гавайская, 21 — Новокаледонская, 22 — Меланизийско-микронезийская, 23 — Полинезийская; 24 — Карибская, 25 — Венесуэльско-гвианская, 26 — Амазонская, 27 — Южная бразильская, 28 — Андская, 29 — Пампасская, 30 — о. Хуан-Фернандес; 31 — Капская; 32 — Северная и Восточная Австралийская, 33 — Юго-западная австралийская, 34 — Центральноавстралийская; 35 — Новозеландская, 36 — Патагонская, 37 — Южных океанических островов.
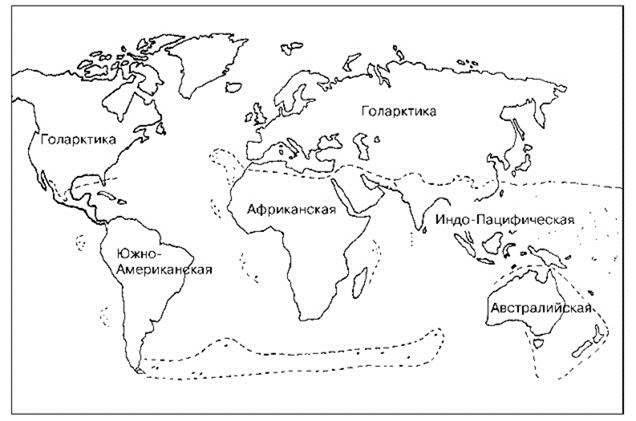
Рис. 3.32. Схема флористического районирования суши Cox (2001). Надписаны области.
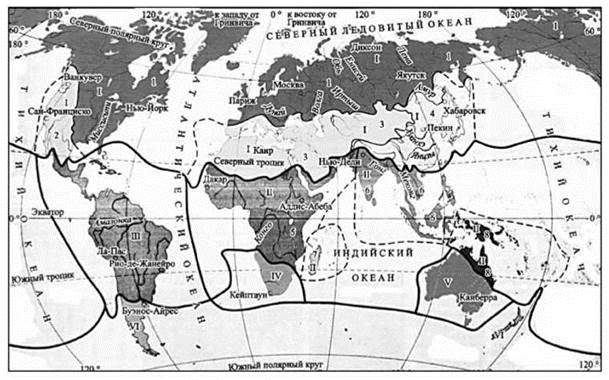
Рис. 3.33. Схема ботнико-географического районирования суши Камелина (2012).
Области (подобласти): I — Голарктическая (1 — Бореальная, 2 — Мадреанская (Сонорская), 3 — Древнесредиземноморская (Тетисовая), 4 — Восточно-Азиатская); II — Палеотропическая (5 — Афро-Аравийская, 6 — Андо-Малезийская, 7 — Мадагаскарская, 8 — Древнеокеаническая); III — Неотропическая; IV — Южноафриканская; V — Австралийская; VI — Голантарктическая.
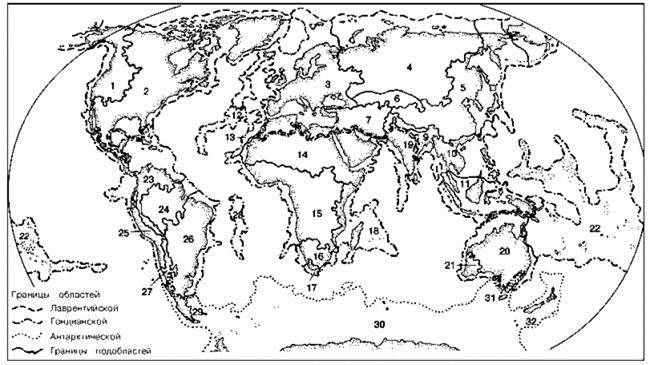
Рис. 3.34. Схема флоро-генетического районирования суши Разумовского (1999).
Области (провинции): Лавразийская (1 — Западно-американская, 2 — Восточно-американская, 3 — Европейская, 4 — Сибирская, 5 — Катазийская, 6 — Джунгарская, 7 — Арианская, 8 — Гималайская, 9 — Бирманская, 10 — Индокитайская, Малайская).
Гондванская (12 — Атлантическая, 13 — Марокканская, 14 — Абиссинская, 15 — Среднеафриканская, 16 — Каффрская, 17 — Капская, 18 — Мальгашская, 19 — Индостанская, 20 — Австралийская, 21 — Западно-австралийская, 22 — Тихоокеанская, 23 — Антильская, 24 — Амазонская, 25 — Перуанская, 26 — Бразильская, 27 — Тукуманская, 28 — Святой Елены).
Антарктическая (29 — Южночилийская, 30 — Кергеленская, 31 — Тасманийская, 32 — Маорийская).
И, наконец, опубликованы интегральные схемы, авторы которых проводят районирование биоты в целом (Рис. 3.35, 3.36).
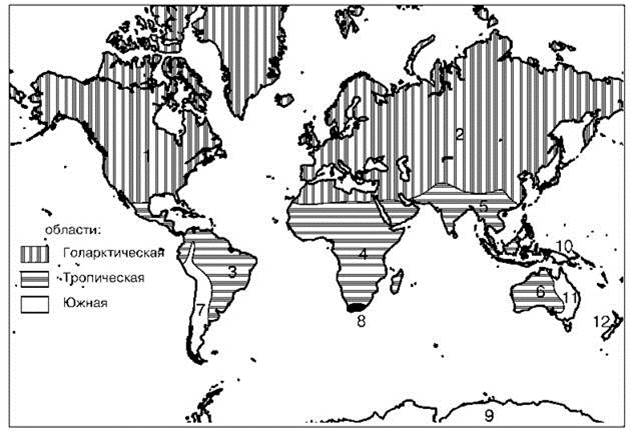
Рис. 3.35. Схема биотического районирования суши Morrone (2002).
Области (подобласти): Голарктическая (= Лавразия) (1 — Неарктическая, 2 — Палеарктическая); Тропическая (= восточная Гондвана) (3 — Неотропическая, 4 — Афротропическая, 5 — Ориентальная, 6 — Австралотропическая); Южная (= западная Гондвана) (7 — Андская; 8 — Капская, или Афроумеренная, 9 — Антарктическая, 10 — Новогвинейская, 11 — Австралоумеренная, 12 — Новозеландская).
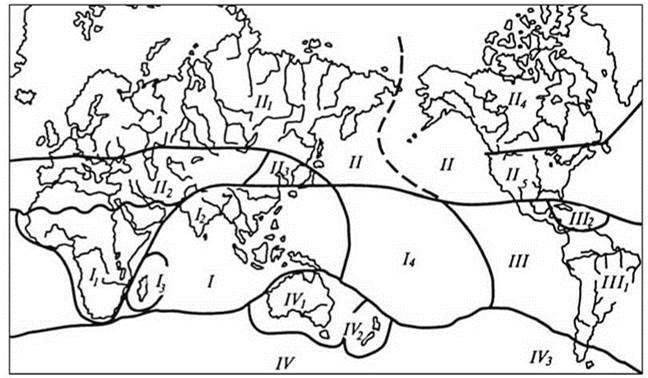
Рис. 3.36. Схема биотического районирования суши Абдурахманова и др. (2003).
Области (подобласти): Арктогея (1 — Европейско-Сибирская, 2 — Древнего Средиземья, 3 — Восточно-Азиатская, 4 — Канадская, 5 — Сонорская), Палеогея (1 — Эфиопская, 2 — Мадагаскарская, 3 — Индо-Малайская, 4 — Полинезийская), Неогея (1 — Неотропическая, 2 — Карибская), Нотогея (1 — Австралийская, 2 — Новозеландская, 3 — Патагонская).
Некоторые территории весьма своеобразны и заслуживают высокого биогеографического ранга. В первую очередь это относится к крупным островам — Мадагаскару и Новой Зеландии. Одни авторы рассматривают их как самостоятельные области, другие — в составе регионов, с которыми они наиболее сходны (Африку в первом случае и Австралию или Юго-восточную Азию во втором). Многие более мелкие длительное время изолированные острова также имеют высокоэндемичные биоты, однако, из-за их малых размеров общее число видов этих биот сравнительно невелико, хотя эндемизм может быть весьма высоким, как на Гавайях, Галапагосах, Новой Каледонии. Он выше во флоре, чем в фауне, особенно в фауне позвоночных, в том числе и потому, что существенная часть островной биоты была уничтожена человеком. Позвоночные были уничтожены в первую очередь, поскольку служили ценным источником белка или представляли угрозу человеку. Поэтому современный эндемизм островных биот меньше, иногда значительно меньше, чем тот, который был присущ их СС до появления человека. Как бы то ни было, мелкие острова принято не выделять в отдельные биогеографические регионы высокого ранга, но, честно говоря, ранг регионов в случае островов довольно субъективен.
Сравнивая разные схемы можно подчеркнуть следующие их основные особенности.
— Фитогеографические карты построены на основе данных по распространению покрыто- и голосеменных растений. Основные их группы имеют мезозойский (юра) возраст. Зоогеографические карты построены преимущественно на основе распространения млекопитающих и птиц, отчасти рептилий и амфибий. Отряды и семейства млекопитающих, птиц и большинства современных рептилий и амфибий возникли в кайнозое. Основные отряды насекомых возникли в мезозое, но многие семейства также имеют кайнозойский возраст. Поэтому часть различий фито- и зоогеографических карт обусловлены разницей в возрасте анализируемых групп, и, соответственно, разной для них возможности к распространению по суше из-за её меняющейся конфигурации.
— Ещё одна из причин отличий фито- и зоогеографии — разная индивидуальная подвижность и продолжительность жизни организмов. Растения неподвижны, но продолжительность жизни деревьев может достигать сотен и даже тысяч лет. Поэтому их стратегия — захват и удержание пространства. Это создаёт структуру наземных СС. Индивидуальные участки даже мелких видов животных обычно охватывают несколько растительных ассоциаций, позволяя им мигрировать в места с более подходящими в каждый момент ресурсами. Это делает связь многих видов животных с растительными ассоциациями менее чёткой, чем видов растений между собой. Особи крупных видов животных могут входить в состав нескольких СС. В результате различия биот крупных животных (позвоночных) разных частей суши часто меньше, чем мелких (беспозвоночных). Эта закономерность видна даже на примере разных отрядов млекопитающих. Таксоны, особи которых крупнее (хищные, хоботные, парнокопытные), имеют почти всесветное распространение, причём некоторые виды имеют огромные ареалы, вплоть до почти всесветного, как у льва Panthera leo, ареал которого охватывал всю Африку, Евразию и северную Америку (Рис. 2.40), тогда как фауны различных частей суши отрядов с более мелкими особями, например, грызунов обладают и гораздо большей специфичностью.
Однако отнюдь не все различия биогеографических схем можно объяснить разным возрастом или биологическими особенностями анализируемых таксонов или различием методов, использованных для их построения. Наиболее ярким примером такого несоответствия является положение границы между азиатской и австралийской биотами (подробнее см. ниже характеристику Австралийской области).
Основа современной биогеографической картины суши была заложена не позднее мела. К этому моменту происходит распад Пангеи и покрытосемянные становятся эдификаторами наземной растительности, происходит становление основных таксонов насекомых. На крупных фрагментах Пангеи возникают разные таксоны наземных животных. Так, в Африке возникли Afrotheria (хоботные, даманы, сирены, трубкозубы, прыгунчики и тенрековые), в Лавразии — Laurasiatheria (насекомоядные, панголины, хищные, непарно- и парнокопытные, китообразные и рукокрылые), в Южной Америке — надотряды неполнозубых и южных копытных.
Последующие изменения топографии суши также наложили свой отпечаток, облегчив миграцию одних и затруднив миграции других. Всё же центры радиации сухопутных таксонов в значительно большей степени совпадают с центрами таксономического обилия тех же таксонов, чем это наблюдается в морском бентосе. Но, как и у бентоса, эти центры не всегда соответствуют месту возникновения таксона.
Принятая в данной книге схема (Рис. 3.37) во многом компромиссна.
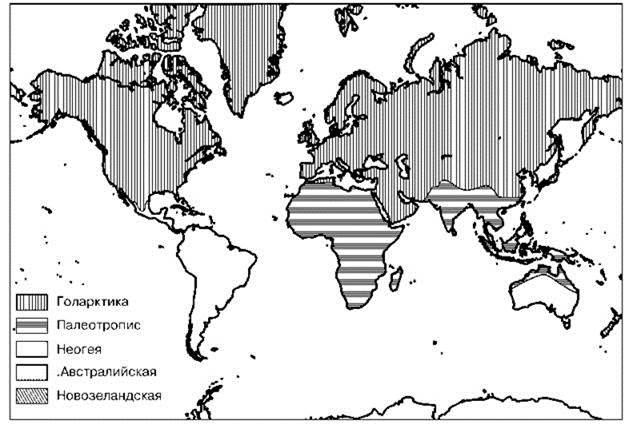
Рис. 3.37. Компилятивная схема биогеографического районирования суши.
Cущественная особенность биогеографии суши состоит в том, что различие схем районирования обусловлено не только различием методов и взглядов их авторов, но и крайне широким распространением антропогенно нарушенных СС. Влияние человека на СС суши значительно больше, нежели на СС континентальных водоёмов, бентали и пелагиали. Это не только вырубка лесов и опустынивание, происходившие и происходящие в историческое время, но и влияние человека на экосистемы Земли в доисторическое время. Поскольку это влияние происходило в ДОисторическое время, биогеографическую картину, сложившуюся к началу ИСТОРИЧЕСКОГО времени воспринимают как нечто естественное. Выше уже было показано, что на всех материках и крупных островах в плейстоцене изменения состояли в первую очередь в исчезновении мегафауны (ниже исчезнувшие таксоны будут описаны) и последующем исчезновении мозаичности СС. Все биогеографы вносят те или иные поправки на антропогенное влияние, но эти поправки очень различны у разных авторов. Практически общепринято, что интродуцированные виды не следует учитывать при биогеографическом районировании, а истреблённые, наоборот, следует включать. Поэтому ниже при анализе состава биот отдельных районов так и сделано. Особенность данного анализа в том, что я попытался не просто учесть истреблённые виды, а максимально учесть изменения СС в результате деятельности человека. Причём не только деятельности Homo sapiens в последние несколько сотен лет, а деятельности разных видов рода Homo за всё время его существования. Результатом этого воздействия было колоссальное изменение в распределении и структуре СС, или, как сказали бы географы, ландшафтов.
По мнению Разумовского (1999) на суше существует всего 174 СС. В то же время в WWF принята система, включающая 14 биомов, восемь областей и 867 экорегионов. Эта схема основана на компиляции региональных схем, построенных самыми разными методами, в том числе и весьма далёкими от биологии. В частности, для территории России основой являлись схема ландшафтного районирования А.Г. Исаченко с соавторами (Исаченко, 1985, Исаченко и др., 1988). Критерием для выделения экорегионов были “во-первых, форма земной поверхности и, во-вторых, растительность” (Olson et al., 2001: 935), поэтому эту схему следует считать географической и она здесь не использована.
Современный уровень знаний не позволяет дать характеристику отдельных СС, поэтому ниже сделана попытка охрактеризовать их объединения ранга области и подобласти. Ниже принято объединение СС суши в четыре области: Голарктика, Палеотропис, Неогея (= Неотропис), Австралия. Обоснование этого дано ниже.
Статус и границы некоторых территорий
Есть несколько регионов, которые одни авторы рассматривают в ранге областей, а другие — нет. Попробуем разобраться в том, кто прав.
Голарктика/Палеарктика/Неоарктика
В биогеографической литературе часто делят Голарктику на евразийскую часть — Палеарктику и североамериканскую часть — Неоарктику (= Неарктику). При этом одни авторы рассматривают их как хорошо обособленные области, тогда как другие — как подобласти одной области, третьи выделяют на территории Голарктики несколько регионов равного ранга, границы которых не соответствуют границе материков. Рассмотрение евразийской и американской частей Голарктики в ранге отдельных областей возможно только при использовании метода сравнения списков биот для сравнения биот Евразии и Северной Америки. Сравниваемые районы в этом случае выделяют по небиологическому логическому основанию: исключительно по конфигурации суши (а в СССР и по политическому делению, поскольку большая часть территории Палеарктики соответствовала территории СССР). Это нельзя признать приемлемым с позиции биологии. Деление Голарктики на Палеарктику и Неоарктику совершенно игнорирует отсутствие различий биот северных районов, в результате районы, населённые мало различающимися или вовсе идентичными биотами относят к разным областям. Различие тайги (включая тундру) и даже северных районов зоны широколиственных лесов — незначительно, при этом границы ареалов ценофильных видов, которые показывают границы СС отнюдь не соответствуют географическому делению. Например, современные лоси относятся к двум видам, различающимся числом хромосом и морфологическими признаками: европейский лось Alces alces и американский лось A. americanus. Ареал последнего охватывает не только Северную Америку, но и Восточную Сибирь и Дальний Восток (Боескоров, 2001).
Необходимо также учесть, что зональность Голарктики в её нынешнем виде в значительной степени имеет антропогенный характер (подробнее см. ниже), хотя, безусловно, и до кардинальной антропогенной перестройки северные районы биотически отличались от южных. Поэтому здесь принята точка зрения ботаников и некоторых зоологов (Бобринского (1951), Петрова (1966) и Крыжановского (2002) и других о нецелесообразности выделения Палеарктики и Неоарктики в любом ранге как естественных биогеографических регионов.
“Главным образом из-за практических соображений, выражения Палеарктика, т.е. восточная часть Голарктики, и Неоарктика, т. е. западная часть, могут быть удержаны, но не в качестве естественных, а лишь как условные понятия” (Бобринский, 1951: 187).