Биогеография общая и частная: суши, моря и континентальных водоёмов - 2017 год
Бореальная зона, или тайга - Широтная зональность - Биогеография суши
Тайга занимает районы с продолжительностью зимы боле 4 месяцев. Примеры климадиаграмм районов произрастания южной тайги показаны на Рис. 3.24, а северной — на Рис. 3.25. За четыре месяца зимы неморальные леса, даже лишённые листьев, испаряют всю влагу, которую накопили за вегетационный период. У хвойных транспирация зимой в 3—12 раз меньше, чем у неморальных деревьев в безлистном состоянии. Как минимум весь эоцен таёжная биота (бывшая тогда компонентом биоты летнезелёных лесов) Евразии и Америки развивалась как единое целое. Большая часть типично таёжных таксонов имеет евразиатское происхождение, часто наиболее древние таксоны происходят из среднеазиатской горной тайги. Эдификаторы лесных климаксных ассоциаций: ели Picea в северных районах и пихты Abies в южных. В Евразии в северной тайге это — ель европейская Picea abies (= ель сибирская P. obovata), на востоке Сибири сменяющаяся P. ajanensis и Шренка, или тяньшаньской P. schrenkiana (в Гималаях, Тибете, Тянь-Шане). Эдификатор климаксов южной тайги — пихта сибирская Abies sibirica. В Северной Америке их заменяют ель сизая, или канадская Picea glauca, и пихты субальпийская, или горная A. lasiocarpa и бальзамическая A. balsamea. Для одной провинции С.М. Разумовский указывает эдификатором климакса кедровую сосну Pinus cembra(= P. sibirica). Ели, пихты и кедровая сосна входят в состав СС неморальных лесов, образуя там ассоциации, поэтому биоты тайги и неморальных лесов имеют много общих видов. В зоне тайги вместо травяного яруса часто развит травяно-кустарничковый, ниже которого расположен напочвенный покров из мхов и лишайников. Поскольку основные деревья этой зоны вечнозелёные, количество солнечной энергии, достигающей почвы во все сезоны невелико, в отличие от неморальных лесов. Поэтому в тайге отсутствуют весенние эфемероиды, характерные для неморальных лесов, а у многих растений нижнего яруса листья живут или более двух лет (вечнозелёные) или более одного года (зимнезелёные). К первым относятся брусника, мхи, плауны, вороника, вечнозелёный хвощ и многие другие. У зимнезелёных растений листья уходят живыми под снег, начинают вегетировать сразу после его схода и отмирают только после появления новой генерации листьев. Слабо развита и энтомофилия, цветы же белой или почти белой окраски. Для тайги также характерно отсутствие ветра, поэтому анемохорные растения отсутствуют, распространение семян происходит благодаря тому, что они (1) очень мелкие — 10-6—10-5 мг (орхидеи, грушанки, споры плаунов, мхов и папоротников); (2) растения образуют сочные плоды, которые поедают насекомые и птицы (ландыш, майник, купена и др.) и даже человек (брусника, черника, голубика, морошка, малина) или (3) семена разбрасываются при созревании (у кислицы до 3—5 м). Широко распространено вегетативное размножение, растения имеют длинные корневища.
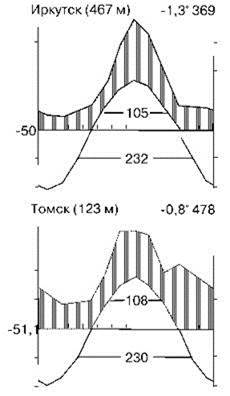
Рис. 3.24. Примеры климадиа- грамм районов южной тайги. По Вальтер (1974).
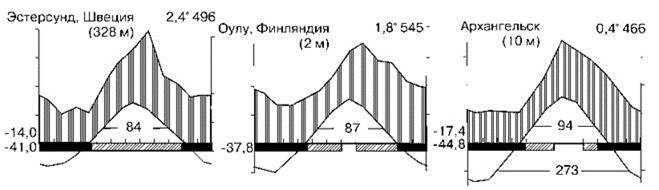
Рис. 3.25. Примеры климадиаграмм районов северной тайги. По Вальтер (1974).
Огромные площади Сибири заняты пирогенными сосновыми борами. Пожары здесь искусственно вызывает местное население, поскольку в пирогенных сосновых борах обильно разрастается ягель — основная пища стад домашнего северного оленя. Возможно, что ещё большие площади лиственичников (не менее 36% российских лесов — лиственичники (Энциклопедия лесного хозяйства, 2006) также имеют сходную природу, поскольку из произрастающих там древесных пород лиственница устойчива к огню даже более чем сосна, лиственница входит в пирогенные сукцессионные ряды в местах её произрастания. Лиственничники расположены в малонаселённых районах, их отсутствие в более населённых местах, вероятно, связано с целенаправленной вырубкой лиственницы, поскольку из произрастающих в Палерактике деревьев она обладает наиболее ценной древесиной, по ценности как стройматериал с ней соперничают только дубы Quercus (граница ареала которых в Европе в голоцене значительно сместилась на юг) и настоящие кедры Cedrus (последние практически полностью были вырублены ещё в Древнем Мире). Например, в Ветлужском ботанико-географическом районе лиственница входит в пирогенный демутационный ряд климакса (Разумовский и др., 1984), но, несмотря на то, что обширные пожары там встречаются чаще, чем раз в 100 лет, даже отдельные крупные лиственницы встречаются только в местах, откуда их невозможно вывезти (крутые склоны оврагов, парки), а лиственничники полностью отсутствуют (как и дубравы — климаксная ассоциация этого района). Из-за большого удельного веса древесины, сплавлять срубленные лиственницы по воде невозможно, транспортировка по суше дорога, поэтому лиственничники и развиты только там, где потребление на месте незначительно. Кедровые сосны Pinus cembra вырублены в том же Ветлужском ботанико-географическом районе настолько давно, что в литературе даже отсутствуют сведения о том, что они там росли, хотя старые деревья встречались ещё в ХХ в. (наши данные). Та же закономерность свойственна и расположенному севернее Сухонскому району (занимает части Костромской и Вологодской областей), где произрастание кедровых сосен отмечено в литературе в XIX в. Несомненно, соотношение антропогенных и природных факторов различно в разных районах, а вырубка деревьев селективна.
При чтении литературы создаётся впечатление, что американские виды хвойных существенно крупнее евразийских. В реальности это в значительной степени это объясняется полным отсутствием в Евразии лесов того же возраста, что и в Северной Америке. Например, в конце XIX в. в Финляндии ещё находили ели Picea abies возрастом 1000 и более лет, ныне такие ели уже не найти. Ежегодный прирост ели и пихты в высоту может достигать 1 м, а лиственницы 1,5 м (наши данные). Ели и пихты возраста 60 лет достигают 30 м высоты, после чего их вырубают. В Северной же Америке большие площади лесов были заповеданы до того, как были вырублены. Вполне вероятно, что евразийские деревья достигли бы сопоставимых с североамериканскими видами размеров, если бы им дали такую возможность.
“Безлесие тундры “обусловлено всем комплексом экологических условий тундровой зоны, режим которой неблагоприятен для развития древесных пород” (Сочава, Городков, 1956). “Эта формулировка, как и все суждения этого типа, означает не решение проблемы, а признание ее неразрешимости и снятие её с обсуждения” (Разумовский, 1999: 93).
Тундра. Безлесие тундры объясняют низкими температурами, рельефом, заболоченностью. Однако, как писал С.М. Разумовский, “природа влажных тундр становится совершенно ясной при ознакомлении с конкретными геоботаническими описаниями. В подавляющем большинстве случаев это ассоциации, образующие в тайге (да и в зоне летнезеленых лесов) стадии гидросерий, причем стадии настолько ранние, что им вообще несвойствен древесный ярус. Поэтому нет никакой необходимости искать специфические для тундровой полосы факторы, делающие невозможным возобновление древесных пород на фоне этих сообществ... Для каждого, кто хоть немного работал в тайге, покажется достаточно убедительным уже простой перечень господствующих мхов, встречающихся в любом описании равнинных влажных тундр: Aulacomnium palustre, Polytrichum commune, P. strictum, Camptothecium trichoides, Calliergon giganteum, Drepanocladus exannulatus, Sphagnum balticum, S. girgensohnii, S. warnstorfii и т.п. В тайге едва ли кто-нибудь удивится безлесности участков, покрытых подобным моховым ковром.
Весьма велика и флористическая общность тундровой полосы и лежащих к югу от нее (в Северном полушарии) таежных пространств. Если исключить ценофобные виды, тундра лишена эндемиков, а ее флора отличается от флоры северной тайги лишь выпадением значительной группы видов, свойственных в тайге зрелым местообитаниям. Все виды, кажущиеся характерными для тундры, глубоко проникают в тайгу по незрелым (главным образом гидросериальным) местообитаниям, хотя и не играют там сколько-нибудь значительной роли в ландшафте; напротив, очень многие таежные виды встречаются в тундре, но, в свою очередь, малозаметны там, будучи приурочены к нетипичным для тундры (т. е. не откровенно болотным) местообитаниям.
Резюмировать все сказанное можно следующим образом. Тундра представляет собой территорию, на которой благодаря весьма высокому коэффициенту увлажнения, равнинности и низменности рельефа, а также близости горизонта вечной мерзлоты преобладают по площади сообщества ранних стадий гидросерий, по самой своей природе лишенные древесного яруса” (Разумовский, 1999: 94—95).
Возобновление леса при достижении сукцессиями древесных стадий затрудняет отсутствие семян деревьев. Поэтому оно происходит только по южной границе тундры. Это выражается в постепенном облесении тундры, наблюдаемом в настоящее время практически повсеместно. Восстановление леса замедляет вырубание деревьев на топливо и поедание древесных всходов стадами северного оленя.
В 1977 г. на научной станции в поселке Черский республики Саха (Якутия) (68° 44’N, 161° 23’E) на территории 16 км2 была начата работа по проекту Плейстоценового парка — воссозданию плейстоценовых СС (официальный сайт проекта http://www.pleistocenepark.ru/ru/). В настоящее время на огороженной территории там живут лошади, лоси, овцебыки, северные олени, маралы и зубры. Предварительные данные показывают, что воздействие мегафауны принципиально меняет облик ландшафта, на смену мхам и лишайникам приходят луга высокопродуктивных однодольных. Похожие проекты находятся на разных начальных стадиях в белорусской части зоны отчуждения Чернобыльской АЭС (в украинской заповедный режим отсутствует), Оренбургской тарпании, или Орловской степи (165 км2 на месте военного полигона), в Объединённых Арабских Эмиратах (о. Сир Бани Яс, 85 км2).
Умеренная зона Южного полушария
В Южном полушарии умеренная зона занимает на несколько порядков меньшую площадь, чем в Северном. Из-за их малых размеров и близости океана температура в течение года колеблется незначительно, тогда как количество осадков гораздо больше и распределены по сезонам они более равномерно.
В отличие от Северного полушария, в Южном нет закономерной смены вечнозелёных деревьев на листопадные. Сплошные леса голосеменных в Южном полушарии в настоящее время почти полностью вырублены, поэтому пояс вечнозелёных хвойных не выражен.
В опубликованных посмертно работах С.М. Разумовский (1999) относит к умеренным районы, в которых древесным эдификатором климакса являются разные виды южного бука Nothofagus, однако в его списке районов к таковым отнесены только Патагония и юг Новой Зеландии. Согласно Вальтеру (1975а), однако, климаксные леса Nothofagusраспространены значительно шире. В Австралии, Тасмании и Новой Зеландии пожары, происходящие чаще, чем раз в 350—400 лет приводят к появлению на месте климаксных лесов Nothofagus более ранних сукцессионных стадий, наиболее ранними ксеросериальными являются заросли миртовых и протейных. Эвкалипты же являются эдификаторами предклимаксных ассоциаций. Широкое их распространение связано с искусственными пожарами, которые практиковали ещё австралийские аборигены для охоты на кенгуру. Также и по тихоокеанскому побережью Южной Америки леса Nothofagus встречаются на север до 36°S.
Умеренный пояс Южного полушария люди осваивали в первую очередь, поскольку можно было просто перенести привычную систему хозяйствования из Северного полушария в Южное. В итоге местные СС во многих местах были уничтожены задолго до того, как туда попали учёные и были созданы заповедники. Поэтому нет никаких свидетельств о том, как выглядели естественные СС до появления там человека. В результате приходится восстанавливать их облик, опираясь на сохранившиеся их остатки. О том, как мало сохранилось ясно из следующей цитаты: “ничтожные остатки более или менее естественной растительности <пампы> можно найти лишь в полосе отчуждения железной дороги, по обе стороны автомобильных дорог” (Вальтер, 1975: 162). В Северном полушарии в таких условиях растут преимущественно ценофобы, полагать, что в Южной Америке всё иначе, нет оснований. Догадаться по составу ценофобов, как выглядели СС абсолютно невозможно. Поэтому к имеющимся в литературе сведениям о безлесных частях СС умеренного пояса Южного полушария надо подходить с изрядной долей скептицизма. Так, в Новой Зеландии естественными безлесным типом растительности считали туссоковые злаковники. Однако оказалось, что они растут на месте выжженных человеком лесов (McWethy et al., 2010).
“На волю! — продолжал кричать географ. — В пампасы!” (Ильф, Петров, 1931 “Золотой телёнок”, пит. по 1975: 150).
“Туссоковый злаковник представляет собой шпальную растительность... лес расти здесь не может” (Вальтер, 1975: 181).
“Каждый автор, желая дать разделение земной поверхности на “естественные” области предлагает всякий раз свое новое деление; то на основании какого-либо одного фактора или на основании той или иной группы животных, считая, что именно эта группа является наиболее пригодной для характеристики фаун” (Лаптев, 1940: 65).
* Кактусовые (Cactaceae) обитают (не считая интродукций) только в Америке. Единственное исключение — эпифитный вид Rhipsalis baccifera, обнаруженный в тропической Африке, на Мадагаскаре, Цейлоне, в Индии и Непале. Существуют две теории его расселения. Согласно одной его семена из ягод были разнесены питавшимися ими птицами. Согласно другой, его семена были развезены по миру вместе с ветками омел, поскольку этот вид кактуса вместе с омелой используют для украшения рождественских праздников (под ветками омелы в английской культуре принято целоваться на Рождество).
** Этот термин (от греч. гилейон — лес) впервые использовал Гумбольдт для влажных тропических лесов Амазонки, позднее так стали называть и ТДЛ других континентов.