Биогеография общая и частная: суши, моря и континентальных водоёмов - 2017 год
Центры разнообразия, происхождения и перераспределения - Островная биогеография и экология - Некоторые крупномасштабные закономерности - Общая биогеография
Широкое принятие “араратской” концепции объясняет многочисленные попытки выяснить место происхождения таксона надвидового ранга путём картирования числа видов (или других подчинённых таксонов). Для этого на карту наносят число таксонов*, отмеченных в каком-либо достаточно маленьком районе и проводят изолинии (Рис. 2.41). При этом выявляют места, где видов много, и места, где их мало. При интерпретации необходимо помнить, что такие карты обычно построены по сравнительно небольшому числу точек, поэтому более или менее достоверными на них являются положение пиков и общий тренд. Границу ареала изучаемого таксона, как обычно, проводят с изрядной долей экстраполяции. Что касается положения изолиний на таких картах, то их проводят с гораздо большими экстра- и интерполяциями на основе недоказанного предположения о плавном изменении числа видов в пространстве, т. е. эти линии — не факты, а их интерпретация. Гораздо более корректным было бы нанесение самих данных на карту, но это делают гораздо реже, возможно потому, что картинки получаются не столь красивые (Рис. 2.42).
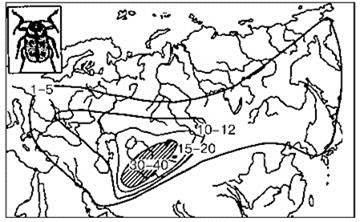
Рис. 2.41. Центры разнообразия жуков рода скрытоглавы азиатские. По Г.М. Абдурахманову и др. (2001).
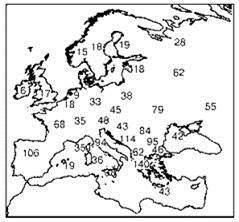
Рис. 2.42. Центры разнообразия покрытосеменных рода смолёвка Silene. По Thompson (1973).
Часто центры таксономического разнообразия интерпретируют как центры происхождения таксона, из которых таксон постепенно расселяется радиально. Такая интерпретация в неявном виде подразумевает два допущения:
1. Периферические виды отличаются по своему возрасту от видов, обитающих в центре (дисперсионистская биогеография).
2. Виды в центре происхождения образуются симпатрически, причём этот способ видообразования резко доминирует в изучаемом таксоне.
Относительно первого допущения имеются две диаметрально противоположные точки зрения. Согласно одной, виды, обитающие в центре происхождения, древнее таксонов, обитающих на периферии. Такого мнения придерживаются основатель кладистики энтомолог W. Hennig и другой энтомолог — W. Brnndin. Согласно другой точке зрения, которой, в частности, придерживается такой известный биогеограф (тоже энтомолог), как Дарлингтон (1966), новообразованные виды вытесняют пред- ковые на периферию ареала. Так же считает и J.C. Briggs (1974,1992, 1995, 2003, 2004). Эта точка зрения согласуется с данными по распределению возраста герматипных кораллов (Рис. 2.43), хотя герматипные кораллы и не являются естественным таксоном, а лишь жизненной формой, да к тому же это является не единственным возможным объяснением уменьшения возраста таксонов из центра разнообразия к периферическим районам. Второе допущение, вроде бы, не очень согласуется с представлением континуалистов о преимущественном происхождении видов путём географической изоляции, хотя, с точки зрения структуралистов, это вполне возможно.
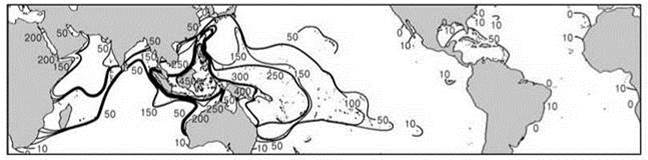
Рис. 2.43. Средний возраст ныне живущих родов мелководных рифообразующих Scleractinia (млн. лет) в разных районах.
Максимальный возраст рода был условно принят за 100 млн. лет, минимальный — 25 млн. лет. По Vernon (2000).
Итак, само по себе наличие повышенного числа видов рода в каком-либо месте может свидетельствовать, а может и не свидетельствовать о месте происхождения данного рода. Поэтому предположения о месте происхождения таксона желательно основывать на палеонтологических данных. Локальное повышение числа видов таксона может лишь привлечь внимание исследователя для последующей интерпретации, не более.
Использование термина “центр происхождения” не только подчеркивает факт повышенного числа таксонов в данном месте (что легко определить), но и утверждает, что они и произошли здесь (а это доказать гораздо сложнее). Поэтому места повышенного таксономического разнообразия лучше называть не центр происхождения, а центр разнообразия. Часто, особенно в группах, по которым палеонтологические данные скудны или вовсе отсутствуют, приходится строить гипотезы о месте происхождения таксона на основе анализа распространения его современных представителей. При этом надо учитывать, что высокое таксономическое разнообразие ЦС (группы ЦС) может быть обусловлено разными причинами:
— Большим потоком энергии через единицу площади её ареала (высокой продуктивностью ЦС). В этом случае таксономическое разнообразие свойственно многим таксонам.
— Большим размером её ареала. В этом случае таксономическое разнообразие свойственно многим таксонам.
— Сильной её фрагментацией. В этом случае таксономическое разнообразие свойственно таксонам с низкой способностью к расселению и/или приуроченным к редким в ареале ЦС биотопам.
— Сильной её изоляцией. В этом случае вспышку таксономического разнообразия дают только некоторые таксоны, при этом многие таксоны, свойственные аналогичным ЦС, отсутствуют.
— Данный регион — рефугиум. Определить это можно по другим группам с хорошими палеонтологическими данными.
А.Н. Миронов (1983, 1985) выдвинул гипотезу, согласно которой повышенное число видов наблюдают не только в центрах происхождения, где выше скорость видообразования, но и в центрах аккумуляции, где более эффективно аккумулируются виды, произошедшие где-то в другом месте. Для характеристики способности района к аккумуляции видов А.Н. Миронов ввёл понятие “таксономической ёмкости района”. Реальность центров аккумуляции он показал на нескольких примерах, два из которых приведены ниже.
1. Анализ фаунистических связей на видовом уровне показывает значительное родство батиальных фаун Индо-Вест-Пацифики и Карибского бассейна. Многие виды, обитающие в этих районах различимы с трудом. Причём большинство видов, распространённых только в этих районах, принадлежат к родам, неизвестным в ископаемом состоянии (по-видимому, они возникли совсем недавно). Причина такого сходства, вероятно, — недавнее формирование батиальной биоты Карибского бассейна. Существовавшая ранее здесь батиальная биота в значительной степени вымерла во время плейстоценовых оледенений из-за понижения продуктивности поверхностных вод (Prell, Hays, 1976) и резкого изменения режима осадконакопления (Emiliani et al., 1975; Be et al., 1976). Факт массового вымирания тропической фауны Западной Атлантики в это время показан для моллюсков (Stanley, Campbell, 1981). Последующее изменение условий в данном районе потребовало формирования новых ЦС, основным источником видов для которых были районы со сходными условиями, а также местные биоты. Индо-вестпацифическая батиальная биота, вероятно, и ранее была наиболее разнообразной, поэтому её вклад в новые ЦС оказался в итоге наиболее существенным. Кроме Карибского бассейна, существенный вклад индо-вестпацифическая батиальная биота сделала и в биоты некоторых других недавно сформировавшихся районов: расположенного в восточной части Тихого океана хребта Наска (Парин и др., 1997) и западноатлантических гор Метеор (Миронов, Крылова, 2006).
2. В миоцене возрос температурный градиент между экватором и полюсами, установилась циркуляция полярного типа и, следовательно, увеличилась продуктивность планктона Северной Пацифики. Одновременно произошло значительное увеличение площади Северной Пацифики (возникли Охотское и Берингово моря). Всё это привело к возникновению обширной абиссальной территории с новыми, ранее не существовавшими в таком масштабе условиями — с богатыми органикой глубоководными илами. На этой территории сформировалась своя ЦС (или несколько). Наиболее преадаптированными к условиям абиссали Северной Пацифики были виды, населявшие антарктическую абиссаль, где аналогичные изменения произошли раньше (циркумантарктическое течение возникло ещё в конце палеогена). Это обусловило наблюдающееся сейчас высокое сходство таксономического состава абиссали Антарктики и Северной Пацифики.
Аккумуляция таксонов при формировании новой ЦС — нормальная и даже обязательная стадия. Вероятно, ещё одним центром аккумуляции была бореальная Западная Атлантика: многие амфибореальные виды, проникшие в Атлантику из Пацифики не смогли закрепиться в Восточной Атлантике (хотя известны в ископаемом виде), но сохранились в гораздо более бедной таксономически Западной Атлантике.
Хотя почти каждый биогеографический район имеет эндемичные виды, некоторые по своему таксономическому разнообразию существенно превышают прочие. Это объясняется характеристиками их ареалов. Возможны два крайних сценария.
1. Если ареал ЦС разбит на ряд изолированных участков (комплекс подводных гор и банок, архипелаг и т. п.), то в некоторых таксонах происходит аллопатрическое образование викарирующих видов в каждом из таких участков. Это даёт эффект повышения таксономического разнообразия за счёт узколокальных эндемов. Такие эндемы занимают очень сходные ниши и любое их число не увеличивает ни сложность ЦС, ни конкурентоспособность входящих в них видов.
2. Если ареал ЦС велик и велик поток энергии через неё, то уже изначально она способна включить в свой состав сравнительно большее число видов. Видообразование здесь происходит симпатрически и вызывается ростом сложности системы в целом благодаря специализации видов.
В реальности мы, конечно, будем иметь дело с промежуточными случаями.
Чтобы подчеркнуть сложность и многостадийность динамики локального таксономического разнообразия, А.Н. Миронов (2006) предложил назвать центры расселения центрами перераспределения.
При формировании новой ЦС в её ареал попадут виды из разных ЦС. Вероятность вхождения видов в новую ЦС будет прямо зависеть от их преадаптированности, их конкурентоспособности и территориальной близости донорской ЦС (близость эту следует оценивать через способность видов к расселению, а не в километрах). Весьма вероятно, что вхождение видов какой-либо ЦС в новую будет значимо повышать шансы вхождения других видов из той же ЦС (поскольку они преадаптированы не только к тем же условиям, но и друг к другу). Виды, входящие в состав ЦС, развивавшихся по второму сценарию, будут иметь при прочих равных более высокие шансы; виды же первых, несмотря на то, что они, казалось бы, также населяют центры разнообразия, будут весьма мало конкурентоспособны. В рассмотренных выше случаях батиаль Индо-Вест-Пацифики и абиссаль Антарктики являются районами, развивавшимися по второму сценарию. Это объясняет существенный вклад их биоты в биоты других районов, даже весьма отдалённых. Виды же из районов, развивавшихся по первому сценарию, так и остаются узколокальными эндемами.
Перемещение ареала таксона или сообщества в пространстве в геологическом масштабе времени называют “прохорез”.