Биогеография общая и частная: суши, моря и континентальных водоёмов - 2017 год
Реликты и рефугиумы - Островная биогеография и экология - Некоторые крупномасштабные закономерности - Общая биогеография
Острова с давних пор являются излюбленным объектом экологов, поскольку их считают удобной моделью для изучения экосистем и эволюции. Во всех учебниках, так или иначе затрагивающих теорию эволюцию, пишут о наблюдениях Чарльза Дарвина за вьюрками на Галапагосских островах. Причина такой популярности островов в первую очередь в чётких границах этих объектов. Наличие чётких границ позволяет абстрагироваться от дискуссии континуалистов и структуралистов, избавляет от тяжёлого и субъективно неприятного труда по проведению границ изучаемых объектов, поэтому кроме островов широкой популярностью пользуются столь же изолированные объекты: озёра, куртины деревьев или кустарников и т. д., причём, чем менее чёткие границы имеет объект, тем менее он популярен. Дополнительными причинами популярности островов являются их постоянные во времени (размер объекта не меняется) и часто малые размеры (легко изучить имеющимися силами), варьирование их размеров и географического положения (имеются легко измеряемые переменные, вдоль градиента которых можно располагать получаемые данные) и некоторые другие. Не последнюю роль, как обычно, играет и мода.
Объекты, которые можно рассматривать как острова, широко распространены в природе. На суше это изолированные горы и горные системы и озёра. Много таких объектов и в море:
— окружающие острова мелководья и их подножия;
— изолированные подводные горы и горные цепи;
— районы гидротерм;
— ультраабиссальные желоба;
— изолированные глубоководные котловины и т. д.
Существенной причиной популярности островной биогеографии является и надежда на практическое применение закономерностей динамики биоты островов в охране природы. Дело в том, что современные охраняемые территории можно рассматривать как острова дикой природы среди антропогенных ландшафтов. Островная биогеография, особенно вначале, казалось, поможет точно определить необходимый размер охраняемых территорий, их размещение и вообще стратегию и тактику охраны природы. Простые закономерности, положенные в основу островной биогеографии, были понятны как политикам, так и их избирателям, что обусловило выделение больших средств на исследования и появление большого количества статей.
Столь большая популярность островной экологии и биогеографии привела к тому, что во многих учебниках и монографиях по общей биогеографии имеются специальные главы, посвящённые ей, и даже публикуются отдельные учебники только по островной биогеографии, например, R.J. Whittaker “Island biogeography”, вышедшая в 1998 г., переизданная в 2001 (дважды), 2002, 2003 гг.
Начало всплеска исследований островной биогеографии было положено Robert H. MacArthur и Edward O. Wilson в 1963 г., когда они опубликовали в журнале “Evolution” статью “An equilibrium theory of insular zoogeography”. Немного позднее, в 1967 г., они опубликовали знаменитую “The theory of island biogeography”.
Теория островной биогеографии в классическом виде — типичная теория континуализма.
Часто число видов на острове прямо связывают с его площадью. Предложены и другие объяснения связи числа видов с площадью:
Гипотеза разнообразия условий. Условия на большей площади более разнообразны, поэтому большее число видов находит подходящие для них условия.
Гипотеза маленьких островов. Маленькие острова из-за своих размеров имеют местообитания, отличающиеся от схожих на крупных.
Гипотеза больших островов. Некоторые виды не могут жить на островах меньше определённого размера из-за недостатка ресурсов на них.
Гипотеза соответствия вида и площади острова. Некоторые виды не могут жить на островах меньше определённого размера, другие — на островах больше определённого размера. Фактически является объединением гипотез 3 и 4.
Гипотеза нарушений. Маленькие острова имеют более неустойчивые местообитания, более подверженные разнообразным нарушениям, элиминирующим большее число видов.
Все перечисленные выше гипотезы предполагают случайное распределение видов. Diamond (1975), изучая распределение видов птиц на островах у Новой Гвинеи, выяснил, что некоторые сочетания видов встречаются гораздо чаще, чем должно было бы быть при случайном заселении острова, тогда как другие — гораздо реже или не встречаются вовсе. Позднее аналогичные данные были получены и другими исследователями. Эти факты не получили простого объяснения в рамках теории континуализма, хотя с позиций структурализма причина такого распределения совершенно очевидна: виды распространяются не независимо, а как члены ассоциаций.
Зависимость числа видов от расстояния до донорской биоты классическая островная биогеография описывает формулой:
St+1 = St+1 - Е,
где: S — число видов на острове в моменты t и t+1,
I — скорость иммиграции, т. е. число видов, прибывающих на остров в единицу времени,
E — скорость вымирания, т. е. число видов, исчезающих из биоты острова в единицу времени.
Графически это можно представить в виде кривых, приведённых на Рис. 2.36 и Рис. 2.37.
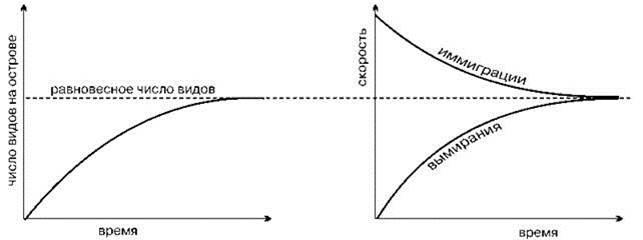
Рис. 2.36. (Слева) Изменение числа видов на острове во времени согласно теории островной биогеографии.
Рис. 2.37. (Справа) Изменение скорости иммиграции и вымирания во времени на острове согласно теории островной биогеографии.
Биологический смысл переменных, входящих в данное уравнение, не столь очевиден, как кажется на первый взгляд. В первую очередь это касается скорости вымирания. Хотя R.H. MacArthur и E.O. Wilson не объясняют причины вымирания, по-видимому, они считают его функцией размера популяции.
Анализ эмпирических данных по динамике видового состава растений, бабочек и птиц на островах Кракатау (Whittaker, 1998) показал, что общее число видов на острове действительно стремится к асимптоте, но по совершенно иным причинам, нежели предсказывает островная биогеография. Скорость иммиграции и скорость вымирания видов меняются не монотонно, что существенно для теории островной биогеографии, а имеет явный пик в начале заселения острова (Рис. 2.38Б). Число видов стабилизируется не в результате установления баланса между иммиграцией и вымиранием, а в результате стабилизации набора видов. Пик в начале заселения острова объясняется, вероятно, первичным формированием ассоциаций, дальнейшая динамика — сукцессионными процессами.
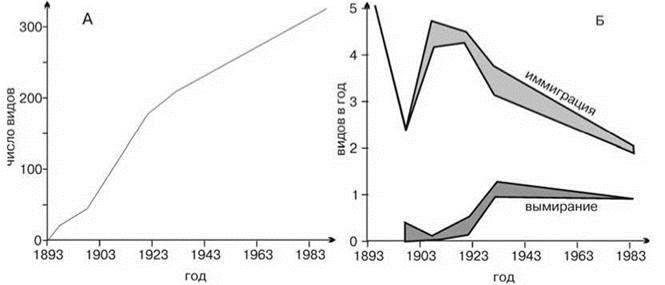
Рис. 2.38. Изменение во времени общего числа видов птиц (а) и скорости их иммиграции и вымирания (б) на островах Кракатау. По Bush, Whittaker (1991) из Whittaker (1998).
В островной биогеографии MacArthur и Wilson (1967) виды принимают идентичными. Между тем, это совсем не так. И это — основная причина того, что, давая красивые результаты на специально подобранных группах островов в пределах одного таксона, данная теория имеет весьма ограниченное применение для предсказаний свойств реальных экосистем.
Предсказания теории островной биогеографии достаточно хороши в тех случаях, когда анализируют острова, являющиеся частью единой ЦС и существующие незначительное время. Но и в таком случае, даже при заселении совершенно новой суши (Кракатау), успешность колонизации острова определяется стадией сукцессионной зрелости покрывающих его в данное время ассоциаций. Безусловно, мозаика ассоциаций имеется всегда, но, тем не менее, вид может прибыть на остров во время (слишком поздно), когда сукцессия уже зашла слишком далеко, или, наоборот, слишком рано, когда ассоциации, в состав которых он входит, ещё не образовались. Своевременность прибытия вида в значительной степени определяет его дальнейшую судьбу на острове.
Методы молекулярной генетики, в частности анализ митохондриальной ДНК, позволяют установить предковые популяции для островных видов. Такой анализ популяций птицы Dendroica petichias на Малых Антильских островах был проведён Klein и Brown (1994). Оказалось, что некоторые острова, по-видимому, были заселены неоднократно. В целом ситуация далека от описываемой stepping-stone гипотезой (популярная теория расселения, в процессе которого вид может образовывать небольшие популяции в маленьких биотопах; как человек, преодолевающий водное препятствие, перепрыгивая по камушкам, такие камушки по-английски называют stepping-stone, отсюда и название гипотезы).
Изучение видообразования на островах и архипелагах во многих случаях показывает вероятность аллопатрического образования близких эндемичных видов. Эти выводы пытаются применить к видообразованию на материках, для чего ищут, а то и постулируют фрагментацию ареалов с последующим их воссоединением. Это увлекательное занятие весьма напоминает популярное в XIX в. строительство мостов между материками для объяснения сходства биот материков, разделённых ныне океанами. Надо отметить, что аллопатрическое образование видов, предполагают, но не доказывают (что, по правде говоря, сделать практически невозможно).
Разница между предсказаниями теории островной биогеографии и фактическими данными может быть обусловлена различной способностью видов к расселению и преодолению преград. Но в ещё большей степени эта разница обусловлена функционированием самих островных ЦС: некоторые условия островов специфичны для них и не имеют аналогов на материках, в частности, это — малая площадь, занимаемая ЦС, и сравнительная однородность условий. Это приводит к тому, что сообщества островов проще, даже если на них не формируется своя ЦС.
Если же на острове (островах) формируется собственная ЦС (т. е. когда он не может быть заселён существующими ЦС и его возраст достаточно велик) предсказания островной теории становятся особенно неудовлетворительными. Это связано с тем, что островная теория в классическом виде не рассматривает видообразование вовсе. Между тем видообразование, обычно сопровождающее формирование ЦС, может принципиально изменить наблюдаемую картину. Возьмём, например, два крупнейших пресноводных озера мира: Виктория и Верхнее. Оз. Виктория имеет максимальную глубину 80 м и площадь в 68 тыс. км2; оз. Верхнее — 393 м и 82,4 тыс. км2. Оба они существуют примерно одно время — около 10 тыс. лет. Однако, оз. Виктория имеет богатую эндемичную биоту (число эндемичных видов составляет многие сотни, велико и число эндемичных родов), а биота оз. Верхнее ничем не отличается от биот соседних озёр.
Островная биота — не случайная выборка из исходной биоты, не группировка. На островах также имеются ЦС. Отличия в их составе от исходных обусловлены различной способностью видов к расселению. Сукцессии могут останавливаться из-за отсутствия видов, необходимых для прохождения данной стадии (диаспорический субклимакс). Одновременно на островах начинается изменение ЦС так, чтобы наиболее эффективно осуществить использование ресурсов при наличном наборе видов. В первую очередь это приводит к расширению экологических ниш видов, часто в сторону ранних сукцессионных стадий (в случае наземных ЦС — это безлесные сообщества).
Изменение ниши видов отражается в изменении их морфологии. В случае наличия большого количества неиспользуемых ресурсов в результате адаптивной радиации могут возникать потребляющие эти ресурсы виды. В результате таксон проходит как бы цикл: виды поздних сукцессионных стадий при формировании новой ЦС или её изменении сначала входят в состав пионерных сообществ, и лишь затем в состав более зрелых, поздних сукцессионных стадий.
“Успешная закладка нового гнезда самкой-основательницей возможна лишь в ограниченном числе мест. Это либо биотопы, непригодные для поселения большинства видов муравьев, либо ситуации “экологического вакуума”, возникающие в результате пожаров, использования химикатов, промышленных разработок, раскорчевки лесосек и других антропогенных воздействий. Именно такие территории оказываются окнами проникновения видов-интродуцентов в новые для них регионы (Passera, 1994)” (Захаров, 2006: 59).
Изучение реальной ситуации в Вест-Индии (Ricklefs, Cox, 1972) показало, что большая часть недавно вселившихся видов действительно несколько расширяют свои ниши, но в целом они приурочены к открытым безлесным сообществам (ранним сукцессионным стадиям), тогда как сравнительно давно вселившиеся виды обитают преимущественно в лесных сообществах (поздних стадиях сукцессии), ниши их сужаются, плотность популяций некоторых из них снижается, других — расширяется (в зависимости от их роли в сообществах). Некоторые виды после вселения сразу занимают узкую нишу и преобразуются в эндемов.
Подобное явление было названо “цикл таксона”. Классическим примером цикла таксона являются результаты исследований R. Wilson (1959), приведённые на Рис. 2.39. В действительности, “цикл таксона” является не какой-либо специфической закономерностью эволюции отдельных таксонов, а отражением становления ЦС — т. е. свойством системы более высокого уровня организации, нежели таксон. Очевидно, что формирование ЦС может начаться только с ранних стадий, а не, например, с климакса. Поэтому термин “цикл таксона” не просто неинформативен, а некорректен и избыточен.
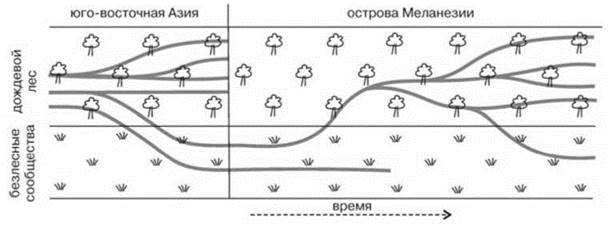
Рис. 2.39. Филогения муравьев Меланезии.
Виды безлесных сообществ при заселении островов сначала входят в состав безлесных сообществ, затем лесных, где образуют несколько видов в результате радиации, позднее от них образуются виды, вновь входящие в состав безлесных сообществ. По Wilson (1959).
Для островных биот по сравнению с материковыми характерны обеднённый видовой состав и упрощённые ЦС, поэтому многие свойства материковых ЦС могут отсутствовать в островных. Степень отличия монотонно возрастает с уменьшением площади, т. е. имеется обратная зависимость между лёгкостью изучения островной ЦС и степенью её отличия от материковых ЦС, изучение которых и является конечной целью большинства работ по исследованию островов. В любом случае, механический перенос выявленных при изучении островных биот закономерностей на материковые невозможен. Более корректным было бы сопоставлять не остров с материком, а ЦС острова (группы островов) с аналогичной ЦС материка. Соответствие теории островной биогеографии фактическому положению вещей тем лучше, чем в большей степени изучаемые острова являются частью крупных ЦС, и тем меньше, чем мельче масштаб, т. е. при переходе с экологического масштаба к биогеографическому точность предсказаний островной теории стремительно снижается. Как вынужден был признать R.J. Whittaker в упомянутой выше “Биогеографии островов”, за более, чем 30 летний период интенсивных исследований выяснилось: “a grand unifying theory remains elusive, and perhaps illusory” (Whittaker, 1998: 4). Поэтому, несколько эпатируя, можно сказать, что биогеография островов — удобная модель для изучения биоты островов. В значительной степени кризис островной биогеографии связан с тем, что её теория возникла на основе идей континуализма, рассматривающих совместную встречаемость видов как результат случайного события. Собранные же данные убедительно показали, что это не так.
С островами связано большое количество и других заблуждений. Так, широко распространено восходящее к наблюдениям Ч. Дарвина мнение о том, что на островах по сравнению с материками гораздо выше доля бескрылых видов насекомых и птиц. Проверка этой закономерности на гораздо большем материале (Raff, 1991, 1994) показала отсутствие достоверного увеличения таких форм по сравнению с материковыми.
С уменьшением площадей, занимаемых отдельными ассоциациями, тенденция к переходу на пастбищные взаимодействия появляется среди групп, у которых на материке такой тенденции нет, например, у мелких грызунов, что приводит к гигантизму островных форм. Это создаёт ещё одну причину для видообразования. Причём она действует даже в тех случаях, когда новая ЦС не образуется, а происходит лишь модификация старой. Но часто “гигантизм” является следствием того, что на необитаемых островах уцелели мелкие представители таксонов, крупные представители которых были истреблены человеком в других местах. Пример такого “гигантизма” — гигантские галапагосские черепахи Geochelone, достигающие 130 см длины и массы 400 кг (Galapago по-испански — черепаха) и вараны острова Комодо Varanus komodoensis, достигающие 3 м длины. В действительности же гигантские черепахи (относившиеся к этому же и другим семействам, до 2,5 м и более длиной) входили в тропические биоценозы плейстоцена по всей суше и исчезали с проникновением в их ареал человека (см. Главу 3). Гигантский варан острова Комодо выглядит карликом по сравнению с австралийским плейстоценовым вараном, достигавшим, по некоторым данным, 8 м длины, и истреблённым человеком. Да и сейчас в южной Азии живёт V. salvator, достигающий 280 см длины, а в Австралии — V. brevicaudata, достигающий 250 см длины. Особи островных популяций могут быть больше предковых материковых или же меньше их. Но их размер определяет не то, что они живут на островах, а оптимизация усилий по добыванию пищи, размера популяции, минимизация пресса хищников и т. п. — но это общие закономерности экологии и эволюции, отнюдь не специфичные для островов.
Биоты изолированных островов формируются из небольшого числа сумевших попасть на них видов. Виды-иммигранты вынуждены как минимум менять ниши, а часто и образовывать другие виды для более эффективного использования имеющихся ресурсов. Так, на Гавайях и в Юго-Восточной Полинезии отсутствуют аборигенные виды муравьёв, это вызвало сильную адаптивную радиацию хищных жужелиц, пауков и даже возникновение хищных гусениц (Paulay, 1994). Процессы, происходящие при этом, уже были разобраны в предыдущей главе при рассмотрении вопросов формирования ЦС.
Реликты и рефугиумы
Особой категорией видов являются реликты (англ. relict от лат. relinquere — оставить позади). Следует различать филогенетические и биогеографические реликты. О филогенетических реликтах говорят в случае таксонов ранга, обычно, не ниже семейства, представленного в данный момент лишь одним (или немногими), но ранее многочисленными видами. Например, Latimeria является единственным современным представителем ранее весьма разнообразной группы кистепёрых рыб, а несколько видов неопилин — немногими современными представителями ранее богатого видами класса моллюсков Monoplacophora. Филогенетические реликты ещё называют живыми ископаемыми.
О биогеографических реликтах говорят в случае, если данная местность ранее входила в сплошной ареал таксона (ЦС), а ныне изолирована от его основного ареала или же произошло резкое уменьшение площади ареала вида. Хорошим примером биогеографического реликта является лев (Panthera leo). Ещё несколько десятков тысяч лет назад ареал вида охватывал Африку, Евразию и Северную Америку. После проникновения человека в разные части ареала лев там более или менее быстро исчезал (подробнее см. Главу 3). Современный ареал льва — жалкие остатки его прежнего ареала (Рис. 2.40).
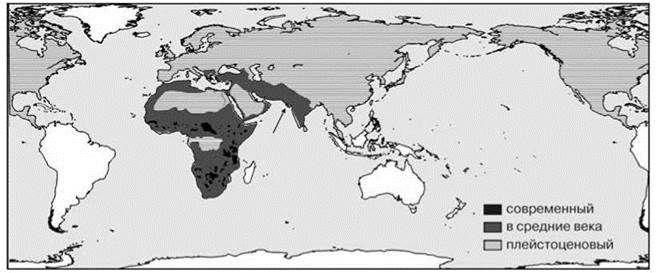
Рис. 2.40. Ареал льва Panthera leo в разное время. Стрелкой показана современная азиатская популяция. Плейстоценовый — по Rothschild, Diedrich (2012), остальные — по Wikipedia.
В 1947 г. филогенетическим и биогеографическим реликтам была посвящена специальная сессия французского биогеографического общества. На ней не только было рекомендовано различать эти понятия, но и предложено использовать для них разные термины. Термин реликт (relicte, англ. эквивалент — relict) было предложено использовать только для биогеографических реликтов, а филогенетические реликты называть relique (англ. эквивалент — relic). Рекомендациям этим, однако, не следуют.
Термин реликт применяют и к надвидовым системам (сообществам, экосистемам). Реликтовые сообщества представляют собой изолированные части ЦС, которые ранее имели в этом районе сплошной ареал, но при изменении климата или по какой-либо иной причине их ареал резко уменьшился. Виды, из которых состоят реликтовые сообщества, если они не входят в состав окружающей ЦС, естественно, будут биогеографическими реликтами. Поскольку сообщества состоят из многих видов, то в занятых реликтовыми сообществами районы сконцентрировано большое число биогеографических реликтов. Такие районы получили название рефугиум (англ. refuge, лат. refugium, мн. число — refugia). Если данные ЦС и виды по-прежнему где-то существуют, а их нахождение в данном конкретном месте является следствием более широкого ареала в прошлом (или результатом перемещения ЦС в пространстве из-за изменений климата), то для таких изолированных участков ареала используют также термин эксклав. Часто биогеографические реликты являются и филогенетическими.
Число обитающих в рефугиуме видов по определению меньше числа видов во всей ЦС (иначе это был бы не рефугиум). Из-за отсутствия соответствующих видов некоторые сукцессионные стадии (ассоциации) невозможны. Поэтому сукцессии в рефугиумах часто завершаются диаспорическими субклимаксами. Устойчивость рефугиумов тем выше, чем более полно представлены в нём сукцессионные ряды, т. е. чем больше в нём ассоциаций. Число ассоциаций проще всего оценить (с определённой ошибкой) по числу видов: чем больше в рефугиуме видов, тем больше в нём ассоциаций. Для устойчивости ЦС наиболее важны блоки, организованные по консортному типу — блоки низших трофических уровней. Блоки высших трофических уровней до определённой степени могут быть заменены другими и тем легче, чем больше в них пастбищных черт.