Биогеография общая и частная: суши, моря и континентальных водоёмов - 2017 год
Субтропическая зона - Основные зоны - Широтная зональность - Биогеография суши
По данным С.М. Разумовского в провинциях Лавразийского царства коренные древесные породы климакса вечнозелёные дубы секции Illex: в Средиземноморье дуб каменный Quercus illex s.lato, восточнее в Гималаях серый, или седой Q. incana, в Японии и Китае дуб убамэ Q. phillyraeoides, в Северной Америке золотистый Q. chrysolepis, сетчатый Q. reticulata, виргинский Q. virginiana, тупой Q. obtusata. В Гондванском царстве коренные древесные породы климакса — лавровые, бобовые и миртовые (к ним принадлежат эвкалипты).
В пределах субтропического пояса выделяют два типа лесов: лавролистные и жёстколистные.
Лавролистные леса (Laurisilvae)
Эдификаторы лесных климаксов высокие деревья с вечнозелёными кожистыми блестящими неопушёнными листьями. Листья ярко-зелёные, простые, цельные, с крупными межклетниками, обращены своей плоской поверхностью к солнцу. Почки защищены почечными чешуями, это отличает их от зональных эдификаторов вечнозелёных тропических лесов.
Растут на Канарских островах (канарский лавр Laurus canarensis), Мадейре, Португалии, тихоокеанском побережье Северной (от Вашингтона до Калифорнии) и Южной (Чили, Патагония с вечнозелёным южным буком Nothophagus betuloides) Америк, Флориде, Японии, южных Гималаях, Новой Зеландии. С.М. Разумовский, опираясь на флористический состав, относит к субтропическим лесам более обширные районы (Рис. 3.21).
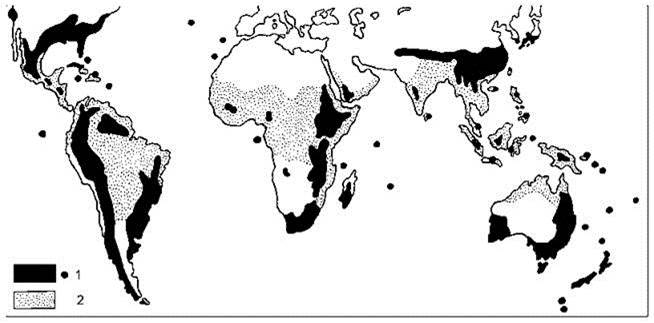
Рис. 3.21. Современное распространение растительности тропиков и влажных субтропиков.
1 — подзона лавролистных субтропических лесов; 2 — зона тропических лесов. По Разумовскому (1999).
Климат равномерно влажный, умеренно теплый. Зональные древесные эдификаторы принадлежат к семействам буковых, лавровых, кунониевых и миртовых. Весьма распространены вторичные и сериальные сообщества хвойных с плоской блестящей хвоей, образующей ветки, напоминающие перистые листья двудольных (секвойя Sequoiasempervirens, тисы Taxus и Cephalotaxus) или с тесносидящими чешуйчатыми хвоями, образующими ветку, напоминающую длинный лист (туи Thuja occidentalis, T. heterophylla).
Жёстколистные леса (Durisilvae)
Древесные зональные эдификаторы отличаются от предыдущих матовыми листьями, сильно укреплёнными механической тканью (чтоб не спадаться во время засушливого сезона), с толстой кутикулой и косым расположением по отношению к свету. Для жёстколистных лесов характерна смена сезонов дождей и засушливого, продолжительность засушливого периода 4—10 месяцев. Количество дождей в общем убывает с удалением от экватора, составляя от 125 мм/год до 1300—1500 мм/год и даже более. Примеры климадиаграмм районов с жестколистными лесами показаны на Рис. 3.22.
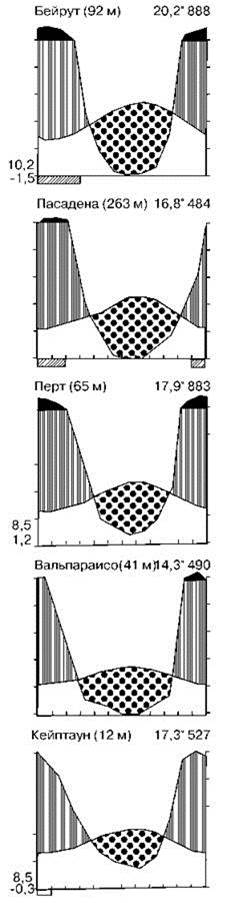
Рис. 3.22. Примеры климадиаграмм районов, занятых жестколистными лесами. По Г. Вальтеру (1974).
Распространены в районах со средиземноморским климатом с сухим и жарким летом и прохладной влажной зимой (см. также Главу 1). Зональные эдификаторы принадлежат к семействам буковых, протейных и миртовых. В Средиземноморье — вечнозелёные дубы Quercus ilex, в Северной Америке разные виды дубов; в Австралии — разные виды эвкалиптов. Лесные климаксы по большей части утрачены и заменены вторичными кустарниковыми сообществами. Безлесные климаксы широко развиты, но часто заменены пустынями, представляющими начальные стадии литосерий.
“Физиономически очень сходные “саванны” занимают большие площади также и более влажных частях Африки и Южной Америки. Сейчас уже нет никакого сомнения, что речь идёт о вторичных растительных сообществах, возникших из различных... типов леса” (Вальтер, 1968: 277).
В низких широтах, в отличие от умеренных, много видов деревьев способны расти отдельно среди травостоя. Поэтому здесь нет чередования лесных и безлесных участков СС, как в умеренных широтах, лишь меняется густота древостоя. Г. Вальтер (1968) указывает, что то, что географы называют саваннами, в действительности — мозаика травянистых и лесных участков (Рис. 3.11). Лишённые же деревьев злаковники, по-видимому, почти всегда антропогенного происхождения, за исключением районов с очень низким количеством осадков, в которых количество осадков слишком мало для произрастания деревьев (Табл. 3.3). В настоящих саваннах деревья просто растут реже, чем в лесу, фактически — это несомкнутые леса с сомкнутым травяным покровом. Поэтому ботаники рассматривают саванны как вариант леса. Географы же, основываясь на современном облике безлесных ландшафтов выделяют их как отдельный тип экосистем (ср. Рис. 3.13, 3.14 и Ц3 с Рис. 3.12 и Ц4). Хотя термин “саванна” взят из языка южноамериканских индейцев, в Америке их называют серрадо, кампос и каатинга (Бразилия), льянос (Венесуэла), чако (Парагвай, Уругвай), мескито (Мексика). В Австралии используют название мэлли. Деревья саванн невысоки (<20 м), среди них много суккулентов. Широко известны стеблевые суккуленты рода баобаб Adansonia: A. digitata в Африке, 6 видов на Мадагаскаре, A. gregorii в Австралии, внешне сходные виды (бутылочные деревья) есть и в Южной Америке. Уже упомянутый эндем острова Сокотра — огуречное дерево Dendrosicyos socotranus (Рис. 1.21) — имеет ту же форму. Подлесок образуют колючие кустарники, в саваннах часто окружающие деревья.
Табл. 3.3. Сравнение аутоэкологических требований деревьев и многолетних злаков. По Вальтеру (1968).
|
деревья |
многолетние злаки |
|
нуждаются в большом количестве осадков осадки могут выпадать зимой запасы воды в почве в засуху должны быть достаточными для минимального расхода растениями высокая влагоёмкость почвы необязательна |
не нуждаются в столь большом количестве осадков осадки должны выпадать в период вегетации (летом) запасы воды в почве в засуху могут быть любыми высокая влагоёмкость почвы обязательна |
Таксономический состав травяного яруса жестколистных лесов сходен на всех материках. Везде основные виды принадлежат к злакам подсемейств просовые Panicoideae и мятликовые, или сорговые Poaceae, обычно крупнодерновинным. В Австралии разнообразие злаков значительно ниже, чем в Африке.
Вторичные кустарниковые сообщества субтропического пояса и примыкающих тропических и умеренных районов В.В. Жерихин (1995) назвал “биомы средиземноморского типа”. Поскольку термин “биом” В.В. Жерихин использовал не в изначальном смысле F. Clements, а как синоним ландшафта, ниже они названы ландшафтами средиземноморского типа (ЛСТ). По мнению В.В. Жерихина, для ЛСТ характерен, прежде всего, низкий уровень использования первичной продукции консументами как непосредственно, так и через детрит. Основная продукция растения создают в холодный и сырой период, неблагоприятный для беспозвоночных — основных фитофагов (второй трофический уровень). В более жаркое время пищевая ценность листьев понижена как из-за увеличения содержания механических тканей, так и из-за накопления специфических метаболитов. Дополнительно фитофагию ограничивает значительная мозаичность растительного покрова, при которой обычно узкоспециализированные фитофаги-беспозвоночные не достигают высокой численности. При антропогенных нарушениях, когда формируются обширные пятна монодоминантной растительности, её потребление интенсифицируется (Fuentes, Etchegaray, 1983). Большинство фитофагов питается не зелёными тканями взрослых растений, а плодами, семенами или всходами, на долю которых приходится лишь малая часть общей фитомассы.
Опад же, наоборот, накапливается в сухой и жаркий сезон, когда недостаток влаги ограничивает активность, как животных-сапрофагов, так и грибов и бактерий, в результате мощность сухой и малопитательной подстилки, на 70—80% состоящей из листового опада быстро возрастает (Read, Mitchell, 1983; Arianoutsou, 1989). Это накопление подстилки не только препятствует возобновлению ассоциаций, но и меняет условия их произрастания, что должно было бы инициировать сукцессию. Этого, однако, не происходит, поскольку для ЛСТ повсюду характерна высокая частота пожаров, не только уничтожающих подстилку, но и воздействующих на растительность. При снижении частоты пожаров они делаются более сильными и разрушительными из-за увеличения запаса горючего материала (Riggan et al., 1988; Minnich, 1989). В результате в районах со средиземноморским климатом (Испания, Португалия, юг Франции, Греция, Калифорния, Австралия) ежегодно происходят катастрофические летние пожары. Значение пожаров для поддержания ЛСТ подтверждает то, что их искусственное исключение в течение 60—100 лет приводит к почти полной остановке возобновления доминантов и существенным изменениям состава растительности (Withers, Ashton, 1977; Recher, Christensen, 1981; Keeley et al., 1986).
Обитатели ЛСТ выработали разнообразные приспособления, связанные с частыми пожарами. В простейшем случае они позволяют переживать пожары: растения имеют запас спящих почек, способны к корневому возобновлению, устойчивы к нагреву стволов и семян, их всходы легко приживаются на открытых участках; животные способны к миграциям (Recher, Christensen, 1981; Shmida, 1982; Mooney, 1983; Zedler, Zammit, 1989). Более сложные адаптации: стимуляция цветения обгоранием и прорастания семян — водными вытяжками из обугленной древесины (Recher, Christensen, 1981; Muooz, Fuentes, 1989; Zedler, Zammit, 1989). Некоторые австралийские и южноафриканские протейные имеют одревесневающие толстостенные плоды, долго сохраняющие жизнеспособные семена и хорошо защищают их, для раскрывания плода и рассеивания семян необходим сильный нагрев (Recher, Christensen, 1981; Mooney, 1983). Аналогично у некоторых хвойных чапарраля шишечные чешуи склеены смолой; семена освобождаются, когда смола растапливается при нагревании (Zedler, Zammit, 1989). У протейных пожарозависимость ещё подчеркивает химический состав семян, богатых азотом и фосфором (которых мало в почвах ЛСТ), но очень бедных кальцием, калием и магнием; успешно прорастают они на свежей золе, обогащённой этими элементами (Stock et al., 1990).
Опад многих растений в ЛСТ обогащён легко воспламеняющимися соединениями, такими как эфирные масла, и не просто допускает, а прямо провоцирует возгорание; это особенно справедливо для эвкалиптовых сообществ.
Свидетельства существования типичного ЛСТ появляются не ранее позднего миоцена-плиоцена (Pons, 1984; Axelrod, 1989). Глубокие адаптации растений ЛСТ к пожарам свидетельствуют о длительном периоде существования пирогенных ассоциаций.
В Средиземноморье в результате деятельности человека “леса были уничтожены полностью... остатки лесов деградировали и уступили место... маквису” (Вальтер, 1975а: 26-27). В Калифорнии “первичный вечнозелёный лес... заменил кустарник, называемый здесь чаппаралем” (там же: 27).
Тем не менее, очевидно, что ряд перечисленных выше свойств ЛСТ имеет вторичную, антропогенную природу. Трудности потребления первичной продукции в дождливый (зимний) период возникают только у беспозвоночных. Крупные растительноядные позвоночные-эдификаторы травяных биомов могли потреблять эту продукцию. В этом случае накопление неразложившегося опада, служащего основным источником горючего материала во время пожаров, должно было происходить в ненарушенных СС гораздо медленнее или вообще не происходить. Уничтожение крупных травоядных животных человеком привело к расширению площадей, занятых пирогенными стадиями ксеросерий существовавших в этих районах СС. Ещё более существенной причиной широкого распространения ЛСТ обусловлено тем, что во многих районах местное население длительно практиковало пожары для целей охоты или скотоводства. В густо населённом Средиземноморье лес уничтожали и непосредственно. Резкое возрастание частоты пожаров и синхронное исчезновение мегафауны, приводящее к такой перестройке растительности, хорошо документировано в Австралии (Rule et al., 2012). Изменение растительности при прекращении пожаров, по-видимому, следует интерпретировать как дальнейшее прохождение сукцессии, остановленной регулярными пожарами, создающими рецидивный субклимакс. Средиземноморский климат соответствует потребностям деревьев, а не многолетних злаков (Табл. 3.3). Настоящим лесным климаксом во всех районах со средиземноморским климатом, как указано выше, являются жёстколистные вечнозелёные леса. Эти леса почти повсюду антропогенно заменены ранними лито- и ксеросериальными ассоциациями. Такие преобразования в разных районах Земли в деталях описаны Г. Вальтером (1968). Поэтому ЛСТ не являются биомами в смысле F.Clements, а лишь районами жестколистных вечнозелёных лесов с сильными антропогенными нарушениями.