Биогеография общая и частная: суши, моря и континентальных водоёмов - 2017 год
Причины эндогенного развития сукцессионных систем - Эволюция сукцессионных систем - Био-экология, или синэкология
Выше мы рассмотрели процесс образования новой ЦС. Её целостность во времени поддерживается постоянством набора ниш и постоянством видового состава. И то, и другое достаточно эффективно регулирует ЦС. Таким образом, ЦС сама по себе, казалось бы, не имеет внутренних причин для развития после того, как она сформировалась. С.М. Разумовский даже считал, что ЦС “или исчезает, или остаётся в неизменном виде. О плавном эволюционировании... не может быть и речи” (Разумовский, 1999: 156). Тем не менее, на протяжении истории Земли одни ЦС сменяли другие, на их смену, в свою очередь, приходили третьи... Основная причина этого, конечно, не в непрерывной бомбардировке поверхности Земли метеоритами. Однако, ни в учебниках экологии, ни в учебниках теории эволюции причины эволюции экосистем обычно вовсе не рассматривают, или ограничиваются общими словами о росте приспособленности видов, полагая, что эволюция экосистем сводится к эволюции входящих в них видов.
Достаточно ясна лишь стадия формирования ЦС. ЦС формируется для более эффективного использования имеющихся ресурсов. Пока находятся неиспользуемые ресурсы, происходит видообразование. При формировании ЦС происходит прогрессивная специализация ассоциаций, сукцессионных рядов и т. д. Это означает рост числа ниш (видов, ассоциаций, сукцессионных рядов и т. п.), плотности их упаковки и уровня коадаптации. Соответственно, при формировании ЦС уменьшается объём ниш видов и сообществ и степень их перекрывания. Однако данный процесс не может продолжаться бесконечно: ни объём используемого ресурса, ни численность популяции, ни площадь, занимаемая ассоциациями, не могут стать сколь угодно малыми. Это объясняет причину резкого замедления темпов эволюции ЦС (и видов) после некоторого этапа формирования, но не помогает понять причину последующих изменений ЦС.
Н.Н. Каландадзе и А.С. Раутиан (1993а,б), анализируя юрский экологический кризис блока наземных тетрапод, выдвинули гипотезу (Рис. 1.53), согласно которой причина эндогенного развития ЦС заключается в следующем. Пока ЦС формируется, направление эволюции видов, входящих в её состав (филогенез), и самой системы (филоценогенез) совпадают. Направление филогенеза — прогрессивная специализация видов. Рост специализации приводит не только к увеличению эффективности потребления видом ресурса, но и к уменьшению потока энергии через популяцию данного вида и к уменьшению его численности, накоплению в системе неиспользуемых ресурсов, а также к росту чувствительности к колебаниям условий его жизни. Поскольку вероятность вымирания любого вида отлична от нуля, рост специализации приводит к увеличению этой вероятности. По достижении некоторого порога степени дробления для сообщества становится невыгодным дальнейшая специализация его членов. Вид же в силу особенностей процесса эволюции стремится к дальнейшей специализации и не может деспециализироваться. В результате дальнейшей специализации видов появляются неиспользуемые ресурсы, которые авторы теории называли “межнишевые лакуны”. Отдельных ресурсов недостаточно для поддержания популяции специалиста, но суммарно разные ресурсы достаточны для поддержания популяции генералиста (неспециализированного вида). Источником генералистов выступают экотоны. Появление в сообществе таких генералистов представляет определённую опасность для сохранения прежней структуры системы. Если структурирующие возможности системы достаточно велики, то на базе этих генералистов возникает новая формация специалистов, в общих чертах воспроизводящая организацию прежней, и устойчивость системы восстанавливается. По мере роста числа специалистов второй формации насыщенность сообщества нишами увеличивается, со временем вновь переходит оптимальный уровень, и цикл повторяется. Модель описывает сообщество, состоящее исключительно из специалистов, причём уровень их специализации примерно одинаков и постоянно растёт. Такое сообщество, если и возможно, отнюдь не является типичным. В сообществах в норме сосуществуют специалисты и генералисты, а уровень специализации видов различен и не может расти бесконечно.
Эту же проблему изучал В.И. Грабовский (2005) на модели, построенной на основе идеологии клеточных автоматов (Рис. 1.54).

Рис. 1.53. Схема сопряжённой эволюции сообщества и биоты, её таксономического и экологического разнообразия.
Типичная последовательность фаз филоценогенеза показана на схеме тремя разными способами в виде: динамики таксономического разнообразия (левая часть схемы); разделения лицензионного пространства и характера преемственности (стрелки) между адаптивными зонами (средняя часть схемы); плотности их упаковки (правая часть схемы). Лицензионное пространство сообщества (1); адаптивные зоны показаны двумя способами: отрезками (средняя часть схемы) или в виде распределений (правая часть схемы) в лицензионном пространстве (2); интерзональные экологические лакуны показаны промежутками между адаптивными зонами (3), нечётко выраженная граница адаптивных зон (4), слабо специализированные таксоны показаны пунктирными линиями и стрелками (5); специализированные таксоны 1-й формации — прерывистыми линиями и стрелками (6), 2-й формации — сплошными линиями и стрелками (7); мера таксономического разнообразия (N); последовательность моментов (T1—T8) геологического времени (7). По Каландадзе, Раутиану (1993а).
В модели рассмотрен простейший случай сообщества, состоящего из одного трофического уровня, виды которого могут специализироваться, и десяти видов ресурсов, пополняемых с постоянной скоростью, т. е. моделировалось сообщество, аналогичное рассмотренному Н.Н. Каландадзе и А.С. Раутианом (1993а,б). Изучение модели показало, что с увеличением потока энергии через сообщество (в природе это происходит при росте продуктивности и/или площади, занятой сообществом), стабильность сообщества растёт. При снижении потока энергии в системе постепенно накапливаются неиспользованные ресурсы. В модели Н.Н. Каландадзе и А.С. Раутиана неиспользуемые ресурсы появлялись в результате сужения ниш видов из-за прогрессивно увеличивающейся их специализации. Согласно их модели, специализация является как бы самоцелью (направление филогенеза — прогрессивная специализация видов). В результате роста специализации входящих в сообщество видов, эффективность использования всем сообществом имеющихся ресурсов снижается, что эквивалентно утверждению о снижении эффективности (полноты) использования имеющихся ресурсов (т. е., в конечном счете, приспособленности) каждым из видов сообщества. Существование такого процесса специализации, который имел бы результатом снижение эффективности использования видом имеющихся ресурсов представляется весьма сомнительным. В модели же В.И. Грабовского накопление ресурсов происходит в результате недостаточно эффективной специализации. Специализация при этом, в противоположность модели Н.Н. Каландадзе и А.С. Раутиана (1993а,б) растёт не безгранично, а только до тех пор, пока растёт эффективность использования ресурсов. Базовые условия модели В.И. Грабовского гораздо больше походят на правду, поскольку специализация, с одной стороны, не может расти неограниченно (как предполагают Н.Н. Каландадзе и А.С. Раутиан), а с другой — всегда недостаточно эффективна. Это происходит из-за того, что (1) непрерывно меняющиеся условия изменяют вектор эволюции и (2) ни одна из ЦС не состоит на 100% из эндемов, какая-то часть видов, зачастую существенная, входит в состав других ЦС. Направление специализации в разных ЦС неизбежно различается, что снижает эффективность использования ресурсов в каждой из ЦС, в которую входит такой вид.
Как только объём неиспользованных ресурсов превысит некоторый порог, сообщество, моделируемое В.И. Грабовским, становится неустойчивым. В нём происходит деспециализация видов на протяжении времени, незначительного по сравнению с периодом стабильного развития. На стадии максимальной деспециализации происходит изменение моды специализации и возникают потребители неиспользованного ресурса. Последние быстро специализируются на потребление неиспользованных ресурсов. Их неограниченное размножение на данном этапе приводит к взрывообразной вспышке численности (этот феномен трудно, если вообще возможно, проследить на палеонтологическом материале). Восстановление баланса производства — потребления приводит в крайних случаях либо к вымиранию новых специалистов, потреблявших ранее непотребляемые ресурсы, и восстановлению старой структуры, либо к установлению нового сообщества со своим характерным заполнением ниш, своеобразным “портретом сообщества”. Обычно, по-видимому, наблюдается промежуточная ситуация. В периоды перестройки эволюция некогерентна, в периоды между перестройками — когерентна. Источником новых специалистов, согласно модели В.И. Грабовского, могут являться деспециализировавшиеся специалисты (в отличие от модели Н.Н. Каландадзе и С.А. Раутиана, где деспециализация считается невозможной). Поэтому перестройка сообщества может происходить только за счёт внутренних ресурсов, использование обитателей экотонов и других сообществ также возможно, но не обязательно даже их наличие. Вероятность деспециализации вовсе не противоречит известному положению о необратимости эволюции, поскольку обратимость эволюции и деспециализация — “две большие разницы”. В возможности деспециализации и последующей специализации к новой нише каждый может легко убедиться, выглянув в окно: большинство синантропных видов прошли именно этот путь.
В модели В.И. Грабовского также представлено, что в устойчивом сообществе одновременно могут существовать и специалисты, и генералисты (на Рис. 1.54 при продуктивности 0,025 и 1) — ситуация, свойственная и природным сообществам.
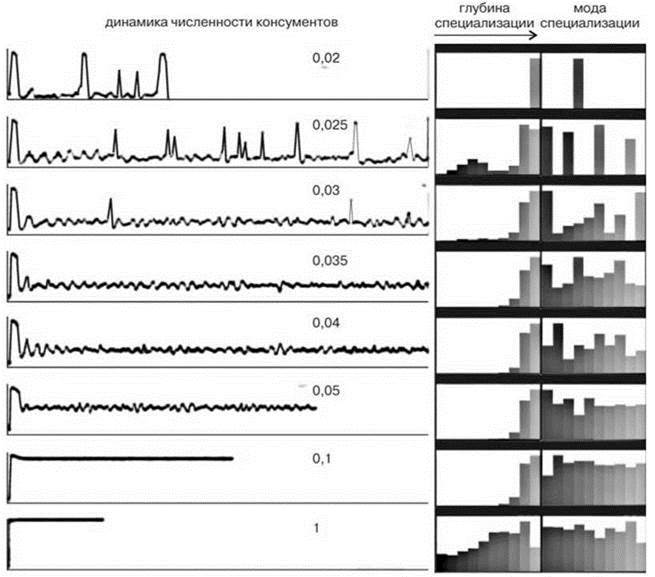
Рис. 1.54. Динамика численности консументов в зависимости от продуктивности.
Цифры — поток энергии через сообщество. Справа: типичные “портреты сообществ”. Глубина специализации имеет 10 градаций; мода специализации отражает потребление 10 различных ресурсов. По Грабовскому (2005).
Данная модель подтверждает, что причины видообразования — экологические.