Биогеография общая и частная: суши, моря и континентальных водоёмов - 2017 год
Океанические биомы - Характеристики отдельных биогеографических регионов - Разные схемы районирования - Биогеография пелагиали Мирового океана
Человечество получает из океана существенную часть продуктов питания, особенно велика роль океана как источника белка. Поэтому необходимо упомянуть о схемах, призванных отражать эту роль океана. Схема районирования FAO приведена на Рис. 6.9. Эта схема принята для сбора статистических данных по вылову различных промысловых объектов, включая и культивированных, она учитывает, как биологические так и прочие данные, границы районов проведены учётом облегчения сбора статистической информации. Более биологической является схема промыслово-географических комплексов Рис. 6.10.
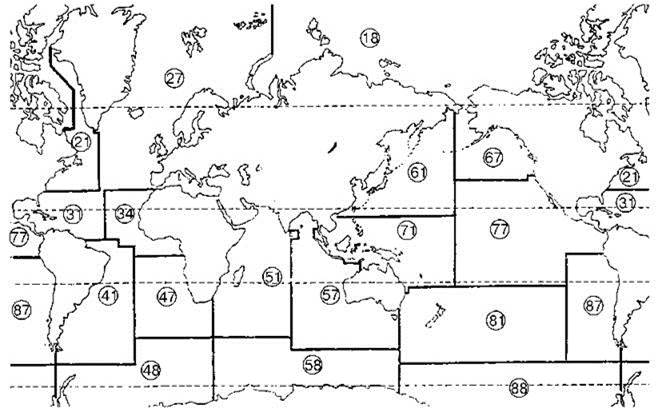
Рис. 6.9. Схема FAO. Основные статистические районы.
18 — Арктические моря, 21 — Северо-Западная Атлантика, 27 — Северо-Восточная Атлантика, 31 — Центрально-Западная Атлантика, 34 — Центрально-Восточная Атлантика, 41 — Юго-Западная Атлантика, 47 — Юго-Восточная Атлантика, 48 — Антарктическая часть Атлантики, 51 — Западная часть Индийского океана, 57 — Восточная часть Индийского океана, 58 — Антарктическая часть Индийского океана, 61 — Северо-Западная часть Тихого океана, 67 — Северо-Восточная часть Тихого океана, 71 — Центрально-Западная часть Тихого океана, 77 — Центрально-Восточная часть Тихого океана, 81 — Юго-Западная часть Тихого океана, 87 — Юго-Восточная часть Тихого океана, 88 — Антарктическая часть Тихого океана.
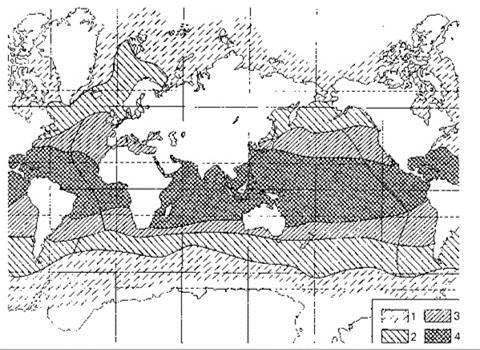
Рис. 6.10. Промыслово-географические комплексы (и районы). По: Расс (1979).
1 — холодноводные (Арктический, Антарктический); 2 — умеренно холодноводные (северобореальные - Атлантический, Тихоокеанский; южнонотальные - Патагонско-Южночилийский, Южно-Новозеландский); 3 — умеренно-тепловодные (южнобореальные - Западно-Атлантический, Восточно-Атлантический, Западно-Тихоокеанский, Восточно-Тихоокеанский; северонотальные в южном полушарии - Перуано-Чилийский, Аргентинский, Южно-Африканский, Южно-Австралийский); 4 — тропическо-экваториальные (Индо-Западнотихоокеанский, Гвинейский или Западно-Африканский, Панамский, Антило-Бразильский).
К.В. Беклемишев (1969) в своей монографии “Экология и биогеография пелагиали”, создавая свою карту районирования пелагиали Мирового океана, основанную на распространении зоопланктона (Рис. 6.12), писал: “для биогеографического деления послужили не ареалы непосредственно, а обобщённые карты основных типов ареалов... все ареалы каждой группы были нанесены на одну карту” (Беклемишев, 1969: 111). Т. е. фактически он использовал метод выделения конкретных биот. Особенности применения метода в данной работе связаны с высокой динамичностью системы течений, в результате которой через границы происходит проникновение не только населения соседних регионов, но и самого биотопа. Из-за больших размеров Мирового океана число проб, собранных в единицу времени на единицу площади невелико, поэтому приходится обрабатывать данные, собранные за дли тельный период времени. Происходившие за этот период перемещения биогеографических границ в пространстве приводят к тому, что на карте отражаются положения границ в разные периоды времени. Фактически это приводит к “размыванию” и без того объективно не очень резкой границы. Поэтому К.В. Беклемишев проводил не одну общую границу, а три: “а) за пределами которой нет ни одного нахождения видов, обладающих ареалом данного типа (“0%”); б) по обе стороны от которой проходит равное число границ ареалов (“50%”); в) в пределах которой встречаются все виды с данным типом ареала (“100%”, или “ядро” данной группы видов)” (Беклемишев, 1969: 111). Использование осреднённых данных приводит к тому, что ареалы отдельных видов, установленные по всем находкам видов, могут существенно отличаться от границ биогеографических регионов. Тот же метод был использован и при районировании пелагиали по фитопланктону (Рис. 6.11), нектонным рыбам (Рис. 6.13) и головоногим моллюскам (Рис. 6.14).
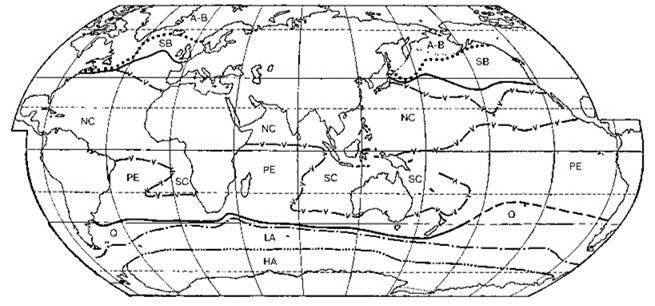
Рис. 6.11. Схема биогеографического районирования по распространению фитопланктона (Беклемишев, Парин, Сёмина, 1977).
А-В — ледово-неритическая зона Аркто-Бореальной области; SB — океаническая зона Аркто-Бореальной области; НА — высокоантарктическая зона Антарктической области; LA — низкоантарктическая зона Антарктической области, включающая Субантарктику; NC — сtверные центральные провинции Тропической области; SC — южные центральные провинции Тропической области; PE — экваториальные провинции Тропической области; Q — флористически обеднённые района неясного ранга на южной периферии тропиков. Тропическая область расположена между сплошными толстыми линиями. В северном полушании флористически обеднённая периферия тропиков не включается в северные центральные провинции.
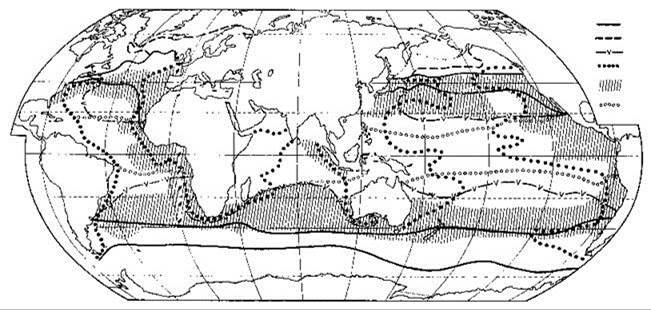
Рис. 6.12. Схема биогеографического районирования по распространению зоопланктона (Беклемишев, 1977).
1 — границы переходных зон; 2 — пределы выселения тропических видов в Бореальной области; 3 — границы распространения экваториальных видов; 4 — границы распространения дальне-неритических видов; 5 — границы распространения периферических видов; 6 — границы между экваториальными и центральными провинциями Тропической области.
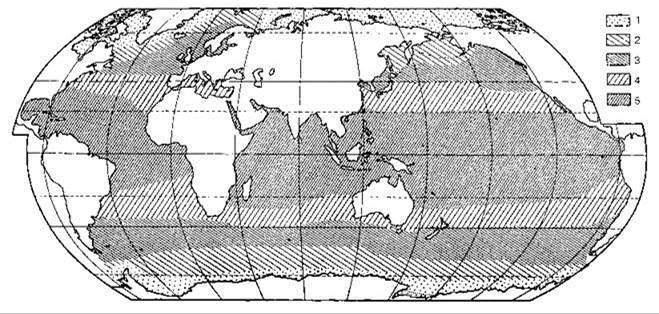
Рис. 6.13. Схема биогеографического районирования по распространению нектонных рыб (Парин, 1968, 1977).
Гомологичные районы обозначены одинаково. 1 — арктический и высокоантарктический; 2 — северобореальные и нижнеантарктический; 3 — южнобореальные и нотальный; 4 — северный и южный субтропические; 5 — тропический.
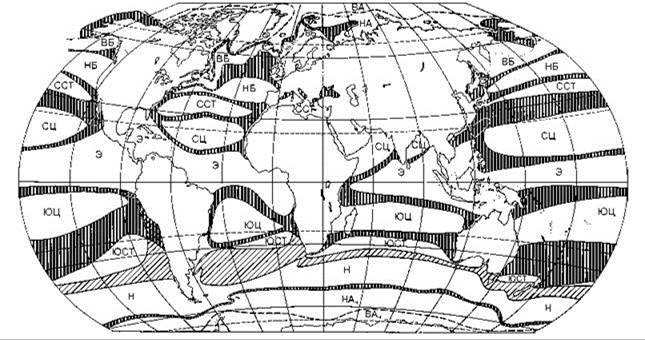
Рис. 6.14. Схема биогеографического районирования по распространению пелагических головоногих моллюсков (Несис, 1968, 1982).
ВА — высокоарктическая, НА — низкоарктическая подзоны арктической зоны; ВБ — высокобореальная, НБ — низкобореальная подзоны бореальной зоны; ССГ — северная субтропическая зона; СЦ — североцентральная, Э — экваториальная, ЮЦ — южноцентральная подзонытропической зоны; ЮСТ — южная субтропическая зона; Н — нотальная зона; НАн — низкоантарктическая, ВАн — высокоантарктическая подзона антарктической зоны. Переходные полосы густозаштрихованы; пунктир — границы подзон арктической и антарктической зон; переходная полоса между южной субтропической и нотальной зонами показана редкой штриховкой.
“Еще 25 лет назад Гентшелю не хватало биогеографических данных для более дробного деления пелагиали в тропиках, и он прибегал к экологическим признакам” (Беклемишев, 1969: 102).
Авторы схем, основанных на распространении нектона, использовали метод типизации ареалов, причём нерестовой части ареала в большинстве случаев придавали больший вес, нежели нагульной. Исключения связаны с особенностями биологии: так, например, у морских лососёвых естественно, учитывают только нагульную часть ареала, поскольку нерестовая находится в пресных водах.
Схема по нектонным рыбам (Рис. 6.13) несколько отличается от схем по фито- и зоопланктону:
— отсутствуют какие-либо зоны со смешанным населением (аналогичные переходным), это связано со способностью организмов нектона активно изменять своё положение в пространстве, при этом виды нектона, входящие в разные ЦС расходятся в пространстве, естественно, более чётко.
— нет экваториальных и центральных видов нектона, по-видимому, это связано с недостатком ресурсов в этих районах.
— нет периферических видов, но зато имеется группа субтропических видов, правда незначительная по числу видов.
А. Longhurst (1998, 2007) акцентировал внимание на том, что, поскольку положение границ биогеографических регионов в пелагиали весьма изменчиво во времени, из практических соображений следует искать способы получения мгновенных “портретов”. В последние десятилетия были созданы новые методы изучения верхних слоёв воды. Наиболее подходящим является изучение данных, полученных со спутников: течений, динамики температуры и концентрации хлорофилла. Такие данные начали собирать с конца 1970-х гг. Это дало исследователям огромное количество новой информации, которая позволяет понять многое в том, как функционируют экосистемы пелагиали и, особенно, их динамику во времени и пространстве. На основе анализа этих, а также традиционных данных А. Longhurst (1998) выделил 54 биома, которые он сгруппировал в два основных типа биомов: океанические и неритические. Океанические, в зависимости от природы пикноклина стабилизирующего фотосинтезирующий слой и динамики вертикальной структуры поверхностной водной массы он разделил на биомы торговых ветров (пассатов), западных ветров и полярные. Биом в понимании A. Longhurst вполне соответствует изначальному смыслу этого термина и используемому в данной книге термину ЦС.
“Важное различие планктонной и нектонной зоогеографии — отсутствие каких-либо зон распространения активноплавающих рыб, аналогичных переходным зонам, выделяемым по планктону и характеризующимся смашанным населением” (Парин, Расс, 1967: 138).
Сравнение схем районирования A. Longhurst (1998, 2007) (Рис. 6.15) со схемами районирования на Рис. 6.11 — 6.14) показывает их значительное сходство. Различия при детальном анализе оказываются скорее методическими. Границами биомов, выделенных Longhurst, являются полярные фронты и субтропические конвергенции (см. схемы в: Бурков, Булатов, Нейман, 1973; Бурков, 1980), что, впрочем и неудивительно. Практически те же границы имеют и зоны, выделенные К.Н. Несисом (1982, 1985), хотя они не вполне гомологичны зонам A. Longhurst. Так, южной границей северной субтропической зоны и северной границей южной субтропической зоны на схеме К.Н. Несиса является северный полярный фронт в Тихом океане и субтропическая конвергенция в Атлантике. Переходная полоса между южной субтропической и нотальными зонами (имеющаяся в схемах Г.И. Сёминой и К.В. Беклемишева) лежит между южной субтропической конвергенцией и южным полярным фронтом и полностью соответствует нотальной зоне Н.В. Парина. Но исследования планктона этого района Н.М. Ворониной (1969, 1984) показали, что в этой “переходной полосе” обитает большое число эндемичных видов, что требует признания за ней статуса самостоятельного биогеографического выдела. Учитывая высокое сходство биогеографических схем, за основу принята экологически более проработанная схема A. Longhurst, откуда заимствованы многие данные по характеристике биомов (ссылки на его работы большей частью опущены, чтобы не загромождать текст). В целом все указанные схемы довольно хорошо соответствуют распределению температурных поясов и системы океанических течений, гораздо лучше, чем это наблюдается для бентоса.
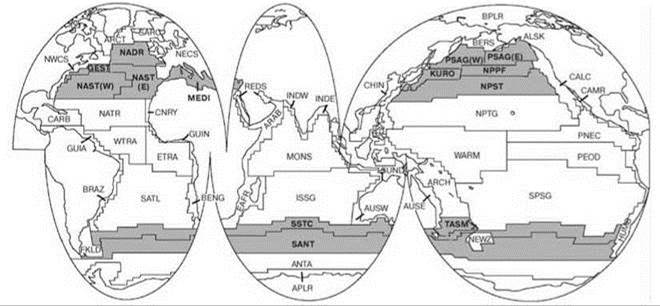
Рис. 6.15. Схема биогеографического районирования пелагиали. По Longhurst (2007).
Полярный биом. Провинции: ANTA — Антарктическая; APLR — Южная полярная; ARCT — Атлантическая бореальная; BERS — Северопацифическая эпиконтинентальная; BPLR — Арктическая полярная; SARC — Атлантическая субарктическая;
Биом западных ветров. Провинции: GFST — Гольфстрима; KURO — течения Куросио; MEDI — Средиземного и Чёрного морей; NADR — Северо-Атлантического дрифта; NAST (E) — Северо-Атлантического субтропического течения (Восточная); NAST (W) — Северо-Атлантического субтропического течения (Западная); NPPF — Северотихоокеанская полярного фронта; NPST — Северотихоокеанская субтропическая; PSAG(E) PSAG(W) — Тихоокеанского субарктического течения (Восточная и Западная); SANT — Южного субантарктического круговорота; SSTC — Южной субтропической конвергенции; TASM — Тасманского моря.
Биом пассатов (торговых ветров). Провинции: ARCH — Глубоководных бассейнов архипелагов; CARB — Карибская; ETRA — Восточная тропическая атлантическая; ISSG — Индийского субтропического течения; MONS — Индийского муссона; NATR — Северо-Атлантического тропического течения; NPTG — Северотихоокеанского экваториального течения; PEQD — Тихоокеанской экваториальной дивергенции; PNEC — Северотихоокеанского экваториального противотечения; SATL — Южного тропического атлантического течения; SPSG — Южнотихоокеанская субтропического течения; WARM — Западнотихоокеанского тёплого бассейна; WTRA — Западная тропическая атлантическая.
Неритические биомы. Провинции: ALSK — Аляскинского даунвеллинга; ARAB — Апвеллинга северозапада Арабского моря; AUSE — Восточноавстралийская неритическая; AUSW — Австрало-Индонезийская неритическая; BENG — Бенгельского течения неритическая; BRAZ — Бразильского течения неритическая; CALC — Калифорнийского течения; CAMR — Центральноамериканская неритическая; CHIL — течения Гумбольта; CHIN — Китайского моря; CNRY — Канарского течения неритическая; EAFR — Восточноафриканская неритическая; FKLD — Югозападных атлантических шельфов; GUIA — Гвианская неритическая; GUIN — Гвинейского течения неритическая; INDE — Восточноиндийская неритическая; INDW — Западноиндийская неритическая; NECS — Северовосточных атлантических шельфов; NEWZ — Новозеландская неритическая; NWCS — Северозападных атлантических шельфов; REDS — Красного моря и Арабского залива; SUND — Зундо-Арафурского шельфа;
Примечание. На приведённой в монографии Longhurst (2007) карте биогеографического районирования показано больше биомов, чем описано в тексте, причем в некоторых случаях из текста ясно, что карта неверна. Воиспроизведенная тут карта исправлена в соответствии с текстом монографии.
Характеристики отдельных биогеографических регионов
В биогеографии пелагиали можно выделить четыре группы регионов (биомов): океанические, неритические, прибрежные и глубоководные. Функционирование океанических биомов не связано с дном, неритических — невозможно без дна. Биотоп океанических биомов — океанические круговороты, неритических — водные массы открытого шельфа, прибрежных — локальные прибрежные водные массы. Биотоп глубоководных биомов — глубинные воды, лежащие ниже главного термоклина.
“Экология и биогеография суть не более, чем удобные слова для обозначения перекрывающихся и плохо выраженных дисциплин...” (Glover, 1961: 202).
В пелагиали часто выделяют несколько вертикальных зон: эпипелагиаль, мезопелагиаль, батипелагиаль, абиссопелагиаль. Эпипелагиаль включает эуфотическую зону и приповерхностный пикноклин (= сезонный в умеренных широтах). Мезопелагиаль подстилает эпипелагиаль. Часто мезопелагиаль делят на верхнюю и нижнюю. В верхней доминируют рыбы с зеркальными боками и частично прозрачные, частично красные декаподы. В нижней рыбы не имеют отражающих свет боков, а декаподы полностью красные. И то, и другое связано с падением освещённости с глубиной: внешний вид более глубоководных видов не учитывает возможность освещения организмов, поэтому они фактически имеют чёрную или близкуую к черному окраску (красные лучи проникают на меньшую глубину, нежели остальные лучи видимого спектра). В эпипелагиали много видов, совершающих суточные вертикальные миграции: ночью поднимающихся вверх, а днём опускающихся вниз. Нередко мигрирующие организмы столь обильны и сконцентрированы, что образуют звукорассеивающие слои, образуюющие на экране эхолота как бы второе дно. Мигрирующие виды из верхней мезопелагиали по ночам пересекают поверхностный пикноклин, тогда как большинство мигрантов из нижней мезопелагиали (большей частью крупные виды макропланктона) останавливаются под поверхностным пикноклином. В отдельных случаях, например, рыбы Ceratoscopelus warmingeri вертикальные миграции достигают глубины 1600—1700 м (Angel, 1989). В то же время большинство более мелких видов макропланктона нижней мезопелагиали не мигрирует вовсе. В пределах эпи- и мезопелагиали солнечный свет играет значительную роль в поведении и распределении организмов. Биоты мезо- и эпипелагиали тесно связаны мигрирующими организмами. Разделение пелагиали на эпи- и мезопелагиаль не подтверждает и анализ ареалов видов. Поэтому ниже они рассмотрены как части одних эпипелагических ЦС.
“Географическое распространение неритических пелагических рыб (анчоусы, сельди, сардины, скумбрии, ставридовые и др.) определяется теми же закономерностями, что и распространение придонных шельфовых рыб” (Парин, Расс, 1967: 132).
Большинство авторов (Беклемишев, 1969; Беклемишев, Парин, Сёмина, 1977) указывают, что биогеографическое районирование неритических районов сходно с районированием бентоса, и отлично от биогеографического районирования пелагиали, не проводя, однако, детального сопоставления. Его сделал К.Н. Несис (1982). Он показал, что наиболее крупные выделы в пелагиали и бентали идентичны, тогда как на уровне более мелких подразделений районирование по бентическим видам более детально, чем по пелагическим.
Общее число пелагических биомов невелико. К 54 биомам, выделенным Longhurst (2007) для эпипелагиали (в узком смысле), надо добавить некоторое (пока неизвестное) число глубоководных биомов. Водные массы, располагающиеся между прибрежным фронтом и берегом Longhurst (2007) специально не рассматривал. Вероятно из-за существенной роли в них донных организмов и их личинок их и не следует рассматривать отдельно от бентических (верхнесублиторальных). Общее число пелагических неритических и океанических биомов (без прибрежных) Мирового океана гораздо меньше 100.
Океанические биомы
Океанические биомы Longhurst (1998, 2007) сгруппировал в полярные (polar), западных ветров (westerlies) и тропические (trades, или trade winds, т.е. пассатов), которые можно считать широтной зональностью открытой пелагиали. Ниже даны характеристики этих групп биомов.
Тропические биомы (биомы пассатов)
Положение. Тропические биомы занимают 22% поверхности Мирового океана, их границы примерно совпадают с 30° параллелью в каждом океане.
Гидрология. Сезонные изменения поступления солнечной энергии недостаточны для зимнего охлаждения, поэтому для них характерна постоянная стратификация эпипелагиали на более тёплый поверхностный и более солёный и холодный подповерхностный слои. Поверхностный слой имеет температуру 28—29°С. Годовая разница между наибольшей и наименьшей температурой не превышает один градус. Подповерхностный слой существенно холоднее. В восточных частях океанов на глубине 35—40 м температура падает до 16°С. Лишь в центральных частях субтропических круговоротов, где происходит опускание воды, стратификация выражена плохо.
Таксономическое разнообразие тропических биомов наиболее велико. Ареалы тропических видов обычно циркумтропические, при более узких ареалах, они обычно охватывают или Индийский и Тихий океаны, или Атлантический.
Фитопланктон. В тропических биомах существуют два типа годовой динамики первичной продукции: постоянно низкая и зависящая от муссонов (см. выше). Первичная продукция низка из-за постоянной стратификации, ведущей к постоянному же дефициту биогенов. Поэтому основные продуценты — мелкие водоросли (пикопланктон), дающие до 90% биомассы и до 80% первичной продукции. Уровень первичной продукции в течение года на большей части площади тропических биомов меняется незначительно. Лишь в районах, подверженных действию муссонов есть аналог биологических сезонов Sverdrup’a (= В.Г. Богорова в отечественной литературе). В начале каждого из двух периодов муссонов происходит смена направления ветра на противоположный (это и называют муссоном). В результате направление течений также меняется на противоположное и апвеллинги сменяются на даунвеллинги. Происходит также заглубление пикноклина, хотя и значительно меньшее, чем в биомах западных ветров, всё же пикноклин часто опускается ниже компенсационной глубины. Это приводит к появлению четырёх сезонов в динамике первичной продукции. Наиболее отчётливо муссонная динамика выражена на севере Индийского океана. В тропическом биоме широко развита фиксация азота атмосферы. Основные азотфиксаторы — цианобактерии Trichodesmum spp. (= Oscillatoria), их распространение ограничено сезонной изотермой 20°С. В фиксации азота существенную роль играют некоторые диатомовые (Rhichelia), имеющие эндосимбиотические цианобактерии. Диатомовые образуют плавающие по поверхности маты размером до 1 м и более. Поскольку в фиксации азота участвует белок, содержащий железо, оно становится лимитирующим фактором. Основной источник железа в открытой пелагиали — пыль, приносимая ветрами с материков (Carpenter, 1989). Постоянно низкая первичная продукция способствует развитию разнообразных симбиотрофных связей, таких как пластиды водорослей, функционирующие в Heliozoa после того, как они съедят сами водоросли.
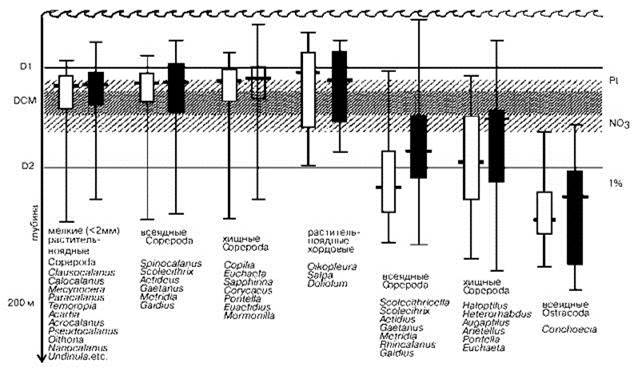
Рис. 6.16. “Типичное” распределение некоторых групп планктона со сходными стратегиями питания в Восточной Пацифике днём и ночью.
Показаны глубины максимальной численности, диапазон глубин 50% популяции и диапазон глубин распределения данной группы за исключением отдельных экстремальных находок. D1 и D2 — границы термоклина, DCM — дневной максимум концентрации хлорофилла, расположен в пределах зоны сравнительно высокой концентрации хлорофилла, Pt — глубина максимальной продукции, NO3 — глубина, на которой нитраты неопределимы, 1% — глубина 1% уровня освещения. По Longhurst (2007).
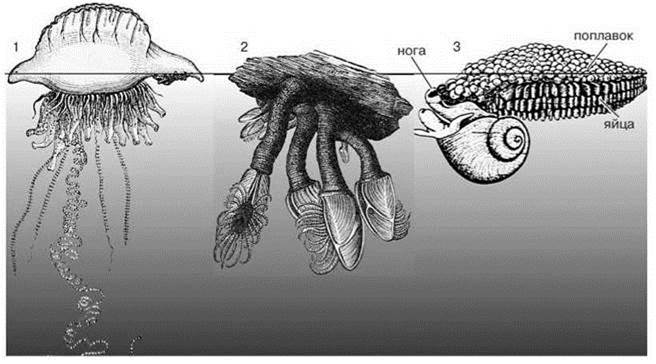
Рис. 6.17. Организмы нейстона.
1 — Physalia (по Перрье); 2 — Lepas (по Перрье); 3 — Janthina (по Руперт, 2010);
Зоопланктон. Стабильность вертикальной гидрологической структуры и равномерное распределение первичной продукции в течение года, позволяет формироваться видам, свойственным отдельным слоям: плейстону, поверхностному и подповерхностному. В результате для низких широт характерно очень высокое таксономическое разнообразие всех групп. Вертикальные миграции мелких фитофагов отсутствуют, редко слаборазвиты суточные миграции. Из-за отсутствия миграций мелких фитофагов более выражены миграции следующего трофического уровня: макропланктона и нектона (кальмары, ракообразные, рыбы). Хотя вертикальные миграции зоопланктона существуют везде, в тропических биомах они особенно отчётливы: существенная часть макропланктона и нектона, особенно крупные копеподы (Pleuromamma, Metridia и Euchaeta), эуфаузиид, светящихся анчоусов Myctophidae и кальмаров поднимается к поверхности вечером и опускается вниз на 200—500 м (до 1500 м) днём. Даже такой крупный представитель планктона, как луна-рыба, питается преимущественно на глубине 110—180 м, затем поднимается к поверхности (0—5 м), где согревается после охлаждения на глубинах (Itsumi et al., 2015). Ежедневные вертикальные миграции фитофагов позволяют фитопланктону за день восстановить свою численность, поэтому фактическая продуктивность в тропических биомах ближе к потенциальной, чем в прочих, где выедание более постоянно во времени (Беклемишев, 1969). Мигрирующий макрозоопланктон и нектон создаёт звукорассеивающие слои.
Роль копепод снижена вдвое по сравнению с полярными биомами (с 68% до 33% биомассы). Соответственно роль других таксонов и общее таксономическое разнообразие повышено, особенно увеличивается роль желетелых: с 1 до 15% и эуфаузиид (с 7,66% в полярных биомах до 30,21% в тропических). Доля хищного зоопланктона от 20—25% в полярных районах возрастает до 40—55% в тропических биомах, около трети этого роста обеспечивают виды, питающиеся желетелыми. Ведущие таксоны зоопланктона — копеподы и эуфаузииды — дают от половины до 70% биомассы (оценка по содержанию углерода). Рыб, питающихся фитопланктоном, нет, по-видимому, из-за очень низкой его концентрации и мелких размеров его клеток. Планктонных чисто эпипелагических рыб мало: лишь карликовые виды макрелещуковых и некоторых других. Гораздо больше планктонных рыб, совершающих вертикальные миграции. Это уже упоминавшиеся светящиеся анчоусы и близкие к ним семейства (Photichthydae, Gonostomatidae)
Нейстон и пелагическое обрастание. Как отмечено выше, настоящие плейстонные ВИДЫ (обитающие в приповерхностном слое воды или на её поверхности постоянно (Рис. 6.18)) есть только в тропических биомах, хотя плейстонные СТАДИИ в развитии видов есть и в других биомах. Положительная плавучесть организмов плейстона обеспечивается газовыми камерами (например, сифонофора физалия) или выделением пенистых поплавков (актиния миниас, гастроподы Jantina, Recluzia, Glaucilla); другие используют как опору поверхностную плёнку воды (например, гастропода Glaucus). Эти организмы плейстона живут непосредственно под поверхностной плёнкой. Усоногие раки Dosima spp., голожаберный моллюск Fiona, их называют гипоплейстон. Другие — на ней (клопы-водомерки Halobates*), их называют эпиплейстон. Третьи частично находятся над поверхностью воды, частично — под ней, как хондрофоры (Velella, Porpita, Porpema), сифонофора Physalia aretusa. В состав плейстона входят даже рыбы: Nomeus — комменсал кишечнополостных, обитающий между их щупальцами, луна-рыба выставляет из воды спинной плавник и использует его как парус. Структуру ассоциаций плейстона создают хондрофоры и Physalia. И те, и другие имеют симбиотические зооксантеллы. Они образуют скопления протяжённостью в сотни миль и плотностью до 1 экз./кв. м и даже более. Их щупальца опускаются на глубину до 10 м и более. Учитывая, что плейстон характерен для олиготрофных районов тропических круговоротов, фактически он — один из существенных продуцентов (Савилов, 1970).
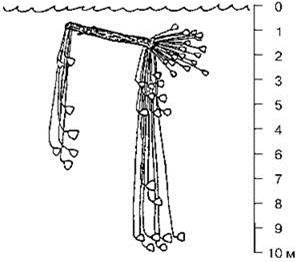
Рис. 6.18. Морские лилии пелагического обрастания мезозоя (на бревне). Hagdorn et al. (2004).
В состав нейстона входят и уже упоминавшиеся маты диатомовых водорослей.
Существенным компонентом нейстона является пелагическое обрастание и оно также имеется только в тропических биомах. Таксономический состав пелагического обрастания весьма специфичен. Биота сообществ пелагического обрастания высокоспециализирована. Генетическая связь между ними и донными сообществами отсутствует. Обрастание это формируется на длительное время плавающих по поверхности брёвнах и других аналогичных с точки зрения биоты предметах (кусках пемзы, а ныне — буях и судах). Несмотря на кажущуюся эфемерность и экзотичность брёвен, судя по тому, что существуют и существовали специфические ассоциации, брёвна являлись и являются постоянным компонентом пелагиали. Специфическими видами, связанными с плавником, являются морские уточки Lepadidae, обычно дающие почти 100% биомассы пелагического обрастания (Рис. 6.18). В другие периоды времени пелагическое обрастание выглядело, по-видимому, совсем иначе. Так, в мезозое в его состав входили колоссальных размеров стебельчатые морские лилии (Рис. 6.19). В пелагическое обрастание входят также саргассовый рыба-клоун Histrio histrio, некоторые рыбы-иглы. Некоторые пелагические рыбы (сайра, “четырёхкрылые” летучие рыбы), как и их неритические родственники, откладывают икру на твёрдом субстрате, в пелагиали таким субстратом является плавник (Парин, 1968). В пелагическом обрастании могут встречаться и неритические виды, имеющие здесь стерильные зоны выселения. Под плавником нередко держатся крупные хищные рыбы.

Рис. 6.19. Рыбы звукорассеивающих слоёв (верхний ряд) и мезопелагиали. По: Парин (1988) и Жизнь животных (1971)
Нектон. Только для тропических биомов свойственны голоэпипелагические рыбы, весь жизненный цикл которых от икры до взрослых особей проходит в эпипелагиали. В более высокие широты некоторые из них заходят летом, когда поверхностный слой воды достаточно прогрет. Большинство видов нектонных кальмаров также ограничено тропиками. Ареалы большинства голоэпипелагических видов циркумтропические, хотя есть и викарирующие индо-тихоокеанские и атлантические виды (Табл. 6.2). Многие виды нектона питаются в мезопелагиали, в частности её обитатели имеют важное значение в питании тунцов, марлинов и меч-рыб (Парин, 1967) и кашалотов. Голоэпипелагические виды нектона имеются только среди высших, филогенетически молодых семейств костистых рыб (преимущественно окунеобразных, несколько десятков видов) и акул, причём голоэпипелагические акулы по палеонтологическим данным известны с начала неогена (Парин, 1968), то же справедливо и в отношении нектонных кальмаров, среди которых активно и хорошо плавающих, всего 6 родов с 1—2 видами каждый (Несис, 1985). Это свидетельствует о весьма недавнем формировании современного нектона. Н.В. Парин (1968) предполагал, что в палеогене была своя голоэпипелагическая биота, но частично она вымерла без следа, и частично была вытеснена в бати- пелагиаль. При этом какие-либо свидетельства того, что палеогеновая голоэпипелагическая биота вообще существовала, отсутствуют, он просто полагает, что коль скоро был биотоп, должны были быть и специализированные виды. Не удивительно, что относительно кальмаров К.Н. Несис (1985) придерживается противоположного мнения: он считает, что после вымирания белемнитов в конце мезозоя — раннем кайнозое, нектонных видов головоногих не было до сравнительно недавнего времени.
“Тропические районы всех океанов почти не различаются по составу ихтиофауны... Тропическая ихтиофауна всех океанов представляет собой вполне монолитное образование” (Парин, Расс, 1967: 135).
В тропических районах имеются специфические формы нектона. Наиболее известны рыбы-лоцманы и рыбы-прилипалы. И те, и другие используют крупных особей нектонных видов почти исключительно для снижения энергетических трат на плавание, а прилипалы — и на дыхание. Многие виды рыб-прилипал связаны с определёнными таксонами или даже видами. Сопровождают крупные объекты для снижения собственных энергозатрат на плавания не только рыбы-лоцманы, но и такие прекрасные пловцы, как тунцы (известны случаи сопровождения ими китовых акул) и дельфины (широко известны факты сопровождения ими кораблей).
Кроме рыб в состав пелагического нектона входят пресмыкающиеся: черепахи и морские змеи (Pelamis platura — неритический вид, могущий обитать и в открытой пелагиали), млекопитающие и птицы. Основной ареал (и места промысла) кашалота Physeter macrocephalus ограничены тропическими биомами.
Стабильность биотопа и сложность биома обеспечивают высокую стабильность высшего трофического уровня — нектона. Например, межгодовые колебания численности рекрутов тропических тунцов на порядок меньше, чем у бореальной трески.
В мезопелагиали постоянно обитает разнообразная фауна (Рис. 6.19). Одни виды постоянно находятся в мезопелагиали и питаются мигрирующими видами, другие, как, например алепизавры Alepisaurus и кинжалозубы Anotopteridae поднимаются к поверхности, где питаются эпипелагическим нектоном.
Биомы западных ветров
Гидрология. Для биомов западных ветров характерно наличие сезонного термоклина. Его появление связано с сезонными изменениями поступления солнечной энергии. Весной уменьшается скорость ветра и увеличивается нагрев поверхности солнцем. В результате стабилизируется верхний слой и появляется разделение эпипелагиали на более тёплый поверхностный и более холодный подповерхностный слои.
Фитопланктон и первичная продукция. В биомах западных ветров существуют два типа годовой динамики первичной продукции: с весенним и зимне-весенним пиками (см. выше). В течение зимы в поверхностной водной массе происходит накопление биогенов, а её вертикальное перемешивание обеспечивает их равномерное распределение по глубине. Поэтому весной в момент появления сезонного термоклина в поверхностном слое биогенов сравнительно много. Это делает возможной весеннюю вспышку фитопланктона. После того, как биогены израсходованы, первичная продукция снижается. Осенью, благодаря заглублению термоклина, возрастанию ветрового перемешивания и опусканию фитофагов на глубину, может происходить осенняя вспышка, но её интенсивность значительно меньше весенней, из-за (1) меньшего количества биогенов, (2) большей толщины поверхностного слоя, (3) меньшей температуры и освещённости. Зимой термоклин разрушается и стратификация эпипелагиали исчезает (наступает гомотермия). Нижняя граница эпипелагиали (в среднем не менее 300 м) расположена значительно глубже компенсационной глубины, что препятствует развитию фитопланктона в нижних её слоях. Также падает общая освещённость и температура. В результате интенсивность фотосинтеза падает, а процессы редукции продолжают идти, это позволяет биогенам постепенно накапливаться, что и обеспечивает весеннее цветение (Рис. 6.20).
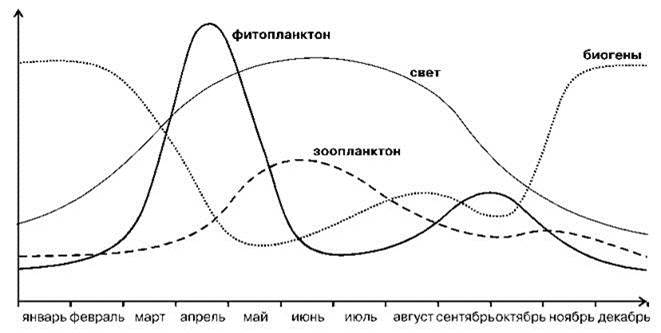
Рис. 6.20. Внутригодовая динамика планктона в Северном море. По Larink, Westheide (2006).
Вышеописанная схема впервые предложена норвежцем Harald Sverdrup в 1953 г. и её считают классической, но в полной мере она описывает только динамику в Северной Атлантике, где исследования начались и были сформулированы первоначальные представления. В других районах вспышки продукции могут происходить даже при отсутствии стабилизации поверхностного слоя. В этих случаях их обеспечивает пикопланктон, обладающей более высокой скоростью размножения. В районах с весенним пиком его ограничивает не только наличие биогенов, но и выедание фитопланктона поднимающимися с глубины фитофагами. В районах с зимне-весенним цветением роль фитофагов незначительна.
Зоопланктон. Крупные виды копепод, особенно в более высоких широтах, осенью опускаются на большие глубины (>500 м) и поднимаются к поверхности ранней весной. Эти миграции, вероятно, являются частью их жизненного цикла, а не обусловлены изменением количества доступной пищи. Для многих крупных видов, по крайней мере в течение части олиготрофной фазы, характерны суточные вертикальные миграции: ночью они находятся у поверхности, а днём опускаются на глубины 200-500 м.
Нектон. Все виды нектона так или иначе, обычно в период размножения, связаны с берегом. Наиболее известны и массовы: сельдь Clupea harengus, откладывающая икру на прибрежную растительность, доминирующая в Северной Атлантике и лососёвые, размножающиеся в пресной воде, Oncorhynchus spp. экологически замещают сельдей в Северной Пацифике. Отдельные виды представлены огромным количеством особей, давая уловы до 2-3 млн. т в год.
Нектонные головоногие представлены только разными видами Gonathus. Более высокий трофический уровень занимают тресковые, морские млекопитающие и птицы. Например, даже в настоящее время в Баренцевом море существенно больше половины нектона и макропланктона (мойва, сайка, сельдь, эуфаузииды и др.), потребляют гренландский тюлень и малый полосатик (Долгов, 2012). Учитывая, что биомасса китообразных и ластоногих существенно снизилась под воздействием промысла, роль млекопитающих в естественных ЦС была выше. В приантарктических водах коммерческий промысел китов, начиная с конца XVIII в. по официальной статистике было убито более 2,7 млн. китов, однако многие страны давали сильно заниженную статистику. Так, СССР занизил свой промысел на 74 000 особей (Brownell, Yablokov, 2002). Многие виды находятся на грани вымирания и, несмотря на десятилетия запрета промысла, пока не показывают признаков восстановления популяций. Китобойный промысел в Северной Атлантике начался гораздо раньше и лишь в Северной Пацифике пик промысла пришёлся на вторую половину XX в., поэтому имеются данные о состоянии ЦС до его начала и на период минимальной численности китов. Произошедшие изменения в структуре пищевых цепей на примере Берингова моря показаны на Рис. 6.21.
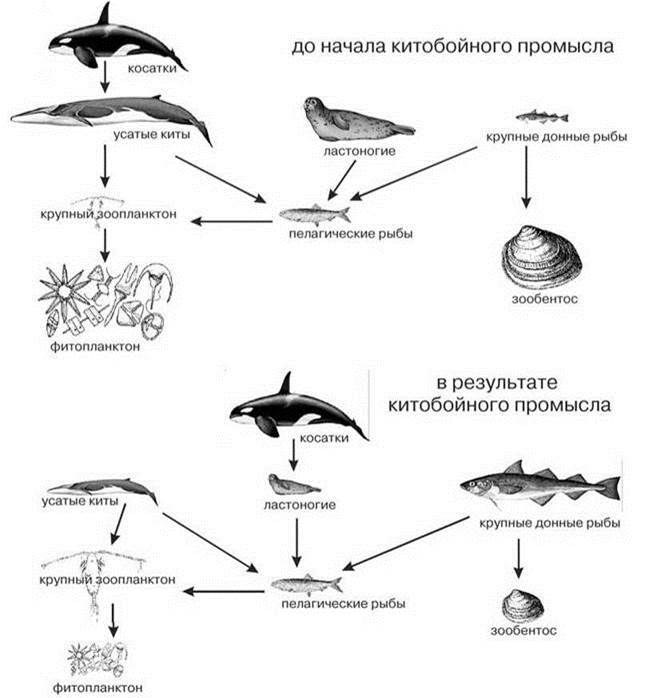
Рис. 6.21. Предполагаемый эффект китобойного промысла на некоторые главные компоненты пелагиали Берингова моря. Функционально доминирующие группы показаны крупным рисунками.
Изменение размеров рисунков отражает увеличение или уменьшение роли соответствующих функциональных групп. До развития китобойного промысла (до 1950 г.) в ЦС доминировали млекопитающие, после (после 1972 г.) — придонные рыбы (минтай, сайда). Уменьшение количества усатых китов привело к увеличению в рационе косаток ластоногих и каланов. Одновременно с интенсификацией китобойного промысла возрос и промысел ластоногих и каланов. Все эти эффекты привели к резкому падению роли в ЦС млекопитающих и росту тресковых. Рост биомассы тресковых (в первую очередь сайды и трески) привели к росту биомассы макрозоопланктона (эуфаузиид и копепод) и, возможно, уменьшению биомассы зообентоса. Рост биомассы зоопланктона может привести к падению биомассы фитопланктона. По Worm et al. (2006).
В Южном полушарии из рыб доминируют нототеноидные, также откладывающие икру на дно. Анадромные виды Северного полушария замещены катадромными (Расс, 1966).
Эпипелагическими видами нектона питаются и мезопелагические хищники. Например в прикамчатских водах около 10% идущих на нерест лососевых имеет травмы, нанесённые мезопелагическими хищниками алепизаврами Alepisaurus и кинжалозубами Anotopteridae (Бугаев, 2015).
Полярные биомы
Полярные биомы характеризуются низким таксономическим разнообразием на всех трофических уровнях.
Положение. Полярные биомы расположены в высоких широтах: они занимают весь Северный Ледовитый океан и часть северо-западной Атлантики, а также воды вокруг Антарктиды внутри полярного фронта. Кроме того, основные черты полярных биомов имеют биомы Берингова и Охотского морей (Северотихоокеанская эпиконтинентальная провинция), поэтому их также следует относить к полярному биому. Полярные биомы Северного полушария расположены в районах, имеющих обширные шельфы, однако, различия акваторий, лежащих над шельфами и вне их невелики, что не позволяет выделить неритические и океанические биомы. В Южном полушарии есть всего два полярных биома, концентрически окружающих Антарктиду, граница между ними соответствует антарктической дивергенции, где происходит опускание холодных вод, создающих глубинные воды Мирового океана, т.е. здесь также невозможно выделить неритические биомы.
Гидрология. Для полярных биомов характерно то, что стабилизацию фотического слоя воды обеспечивает поверхностный распреснённый слой (верхний слой распреснённой линзы). Основные источники пресной воды в Антарктике — тающий лёд, в Северном Полушарии — сток рек и тающий лёд. Поскольку распреснённая линза ограничивает вертикальную циркуляцию, в полярных биомах образуется лёд, по крайней мере, зимой (подробнее см. Главу 5).
Фитопланктон и первичная продукция. 75—90% первичной продукции создают крупные диатомовые и кокколитофори- ды, остальное — пико- и нанофитопланктон. Лёд препятствует ветровому перемешиванию поверхностного слоя и снижает его освещённость: всё это сдерживает развитие фотосинтетиков. Лишь в самой толще льда и на его нижней поверхности формируются специфические сообщества, включающие виды, тесно связанные со льдом — криопелагические (Андрияшев, 1966), или ледово-неритические (Беклемишев, 1969). Разрушение льда приводит к резкому возрастанию освещённости и перемешиванию распреснённого поверхностного слоя ветром. Это приводит к вспышке фитопланктона за счёт крупных диатомовых. Весной кромка льда быстро отступает к полюсам (до 10 км/день), соответственно меняется в пространстве и положение максимальной продукции и биомассы фитопланктона (Рис. 6.22), а вслед за ним — и растительноядного зоопланктона и хищников. Многие виды мигрируют вслед за передвижением кромки льда. В северных полярных районах изначальное содержание биогенов в распреснённой линзе гораздо меньше, чем у Антарктиды. Это приводит к тому, что там фитопланктон обычно выедает биогены полностью, тогда как у Антарктиды это наблюдается редко и только вблизи берегов. Объяснений этому феномену много, но общепринятого нет. Между вспышками после исчезновения биогенов в фитопланктоне доминируют мелкие водоросли. Для Антарктики, где в районе антарктической дивергенции постоянно происходит подъём обогащённых биогенами вод (апвеллинг) высокая первичная продукция создаётся в течение всего года, меняется лишь локализация мест, где она наиболее высока. В Арктике высокие величины первичной продукции характерны только для весенней вспышки фитопланктона.

Рис. 6.22. Схема изменений концентрации хлорофилла А в Баренцевом море (мг/м3). По Wassmann et al. (2006).
Зоопланктон. Непостоянная в течение года вертикальная структура и длительные периоды гомотермии делают невозможным формирование видов, свойственных поверхностным и подповерхностным водам. В результате для умеренных и высоких широт характерно очень низкое таксономическое разнообразие. Основные фитофаги — разные виды рода Calanus и другие копеподы. Для крупных копепод свойственны длительные циклы развития (два и более года) и сезонные вертикальные миграции. В первый год они достигают V копеподитной стадии (предпоследней), после чего мигрируют большие глубины (до 2000—3000 м), где температура постоянно отрицательная или низкая положительная. Это позволяет им снизить расходы на обмен и уменьшить потери от выедания. В период весенней вспышки фитопланктона копеподиты поднимаются в поверхностные воды и завершают жизненный цикл, приступая к размножению. Там, где глубины недостаточны для совершения отногенетических миграций, копеподы не завершают свой жизненный цикл и образуют зависимые популяции. Численность некоторых видов копепод огромна и, вероятно, превышает даже численность наиболее массовых видов наземных насекомых.
В противоположность широко распространённому мнению о большой роли эуфаузиид в зоопланктоне полярных районов, эуфаузииды дают <1% по численности и <8% по биомассе (в пересчёте на углерод) от всего зоопланктона. Особенно мала роль эуфаузиид в Арктике (0,04% по органическому углероду). Но невелика она и в Антарктике: в большинстве других биомов роль эуфаузиид вдвое-втрое выше, чем в Антарктике, в тропических биомах они создают до 30% биомассы. Даже в неритических биомах они иногда тоже могут создавать очень большие скопления. Так А.Г. Погодин с соавторами (1982) описывает уловы эуфаузиид промысловым тралом до ста тонн за одно траление в Татарском проливе. Copepoda высоких и умеренных широт в условиях недостатка пищи опускаются в глубинные слои и впадают в дипаузу, неподвижно вися в толще воды. Низкая температура более глубоких слоёв дополнительно снижает траты на обмен. Нейтральную плавучесть обеспечивают запасы липидов и восковых эфиров, например, у Calanus hyperboreus липиды составляют до 60 % от сухой массы тела (Scott et al., 2000). Эуфаузииды — более крупные животные, размеры которых на порядок превышают размеры копепод. Вес Euphausia superba — около 1 г. Это слишком много для парения в воде: будучи неподвижным, рачок за три часа опускается на 500 м. Для поддержания нейтральной плавучести с помощью регуляции удельного веса тела эуфаузиидам требуются значительно большие запасы липидов, видимо, превышающие 50% от массы тела. Создание таких запасов, скорее всего, в высоких и, вероятно даже умеренных широтах, просто невозможно: содержание липидов у T. raschii в середине зимы в Баренцевом море не превышает 40% от сухой массы тела (Sargent, Falk-Petersen, 1981). Поэтому, чтобы не тонуть, эуфаузииды должны постоянно двигаться и питаться, а при недостатке корма — расходовать внутренние резервы. Это вынуждает их постоянно находиться в поверхностном слое воды с более высокой температурой, приводящей к более высоким тратам на обмен. Метаболизм E. superba, с учетом размеров рачков и температуры воды, необычно высок, что обусловливает потребность в большом количестве пищи. Для её удовлетворения необходимо ежедневно отфильтровывать 50—100 л морской воды и съедать около 35 мг фитопланктона (сырой вес). Поэтому эуфаузииды не совершают вертикальных миграций. По-видимому, именно различиями стратегий переживания неблагоприятных условий можно объяснить то, что копеподы способны образовывать независимые популяции там, где сезон продуцирования органических веществ фитопланктоном непродолжителен по времени и локализован в пространстве (трещины, разводья во льдах, полыньи), например, под многолетними льдами в Центральном Арктическом бассейне. Для эуфаузиид подобные районы являются областями стерильного выселения (Тимофеев, 2005). Более того, эуфаузииды не размножаются регулярно в Арктике (Ломакина, 1964). Поэтому низкая роль эувфаузиид в высоких широтах и, особенно, в Арктике — низкая и очень неравномерно распределённая в течение года первичная продукция в Арктике.
Антарктический криль (эуфаузииды) более чем наполовину сконцентрирован в юго-западной части атлантического сектора. Скопления Euphausia superba в Антарктике при толщине по вертикали около 10 м могут простираться на километры. В них плотность особей составляет до 30000 экз./1 м3 воды. Живой вес такого скопления может достигать многих миллионов тонн. Синий кит, самое большое известное животное, питается в основном эуфаузиидами. Его желудок вмещает 1200 л криля, что соответствует весу более 1 т, а ежедневный рацион — 4 т криля. Кроме китов, крилем питаются тюлени, рыбы и морские птицы (Рис. 6.23).

Рис. 6.23. Схема пищевых цепей Антарктической провинции.
В центре — криль Euphasia superba, его окружают его потребители: тюлень, птица, пингвин, усатый кит, кальмар, нототениевидная рыба и медуза. Рисунок Н.Н. Кондакова.
В Арктике на второе место по биомассе в зоопланктоне вместо эуфаузиид выходят птероподы, дающие 10% С, что существенно выше, чем в любом другом районе Мирового океана (хотя птероподовые илы отсутствуют в Арктике и найдены только на вершинах глубоководных поднятий, верхних частях склонов в тропических и субтропических районах).
Нектон. Таксономическое разнообразие большинства таксонов нектона мало. Неритические рыбы нередко концентрируются у нижней поверхности льда.
Число видов рыб полярного биома Антарктики <1% от мировой морской фауны, среди них нет пелагических видов. Большая часть рыб Антарктики (66—82% в зависимости от источника) принадлежит к Нототениевидным, все они имеют донную икру. В гораздо более молодом северном полярном биоме разнообразие ещё меньше, основным компонентом макропланктона и пищей птиц и млекопитающих является сайка Boreogadus saida и арктическая треска Arctogadus glacialis, а в южных районах — мойва Mallotusvillosus, но икра их пелагическая. Нектонные головоногие представлены только разными видами Gonathus spp. Разнообразие водных млекопитающих (тюленей и китов) в полярных биомах гораздо выше, чем в тропиках, но ниже, чем в биомах западных ветров. Некоторые таксоны морских птиц также полностью или почти полностью эндемичны для полярных биомов, например пингвины — для Антарктики и нотальных вод и замещающие их экологически чистиковые и гагары в Северном полушарии.
Криопелагическая биота. Лёд, как твёрдое тело, выступает некоторым аналогом морского дна. Поэтому в составе населения льда встречаются многие бентосные организмы. Однако основу экосистемы морского льда создают специфические виды.