Биогеография общая и частная: суши, моря и континентальных водоёмов - 2017 год
Фронты - Физико-географическая характеристика Мирового океана - Биогеография бентали Мирового океана
Взаимодействие водных масс друг с другом и с дном приводит к появлению зон повышенных градиентов абиотических факторов — фронтов. С биологических позиций фронты — это экотоны. Можно выделить три основных типа фронтов.
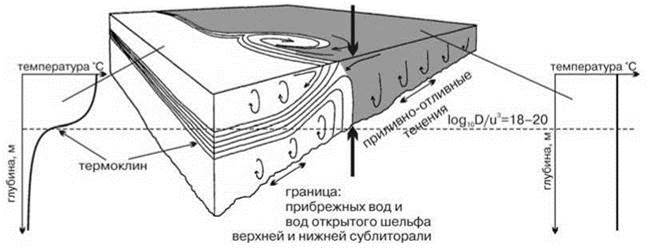
Рис. 5.25. Формирование прибрежного фронта.
Прибрежная водная масса — тёмно-серая. Показано характерное распределение температуры по глубине в прибрежной водной массе и мористее неё. D — глубина, u — скорость течения на поверхности. По: Longhurst (2007)
Прибрежный фронт. Приливы и отливы не только приводят к периодическому затоплению и осушению литорали, но и создают течения. Эти течения, как и любые другие, взаимодействуя с дном, становятся из ламинарных турбулентными. На некоторой глубине D турбулентность эта доходит до поверхности, т. е. распространяется по всей толще воды (Рис. 5.26, Рис. 5.27 Б), поэтому на глубинах, меньших D, вода регулярно один-два раза в сутки перемешивается от поверхности до дна. Штормы, увеличивая скорость течения и создавая ветровое волнение, дополнительно усиливают перемешивание. В результате у берега возникает прибрежная водная масса, для которой характерно (1) отсутствие постоянной стратификации (нет даже сезонного пикноклина), следствием чего является (2) практически непрерывное поступление биогенов в фотический слой и очень высокая первичная продукция. Мористее, на больших глубинах расположена водная масса открытого шельфа, для которой характерен сезонный (в умеренных широтах) или постоянный (в тропиках) пикноклин, затрудняющий поступление биогенов из придонных слоёв. Граница между прибрежной водной массой и расположенной мористее водной массой шельфа (прибрежный фронт), расположенная на глубине D в каждый конкретный момент достаточно хорошо выражена, хотя её положение и непостоянно во времени (Рис. 5.26), поскольку D зависит от скорости течения на поверхности в кубе. Это означает, что:
— D не является константой. Она меняется два раза в месяц (квадратурные и сизигийные приливы); зимой D больше (сильнее ветер и больше скорость течения), но выраженность границы между водными массами меньше, поскольку в открытом шельфе также развивается вертикальная циркуляция из-за охлаждения поверхностных слоёв;
— в местах (и в периоды) с быстрым течением D может быть равна 100 м и более, тогда как в расположенных рядом закрытых бухтах в то же самое время — всего нескольким метрам.
Поэтому при проведении биологических исследований ориентироваться следует не на глубину, а на гидрологическую структуру, которая более отчётлива летом. В открытых районах наиболее часто прибрежный фронт расположен над глубинами 50—100 м. В морях, где приливы не выражены (Чёрное, Японское, Средиземное), граница прибрежной водной массы расположена на гораздо меньших глубинах, чем в приливных.
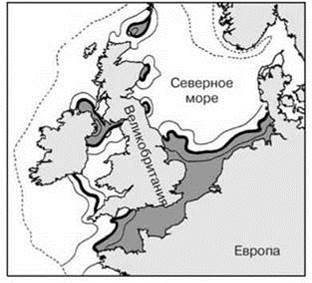
Рис. 5.26. Положение прибрежного фронта у Британских островов.
Толстая линия — среднее положение фронта, тонкие — крайние его положения, пунктир — изобата 200 м. По Longhurst (1998, 2007). Прибрежная водная масса залита тёмно-серым.
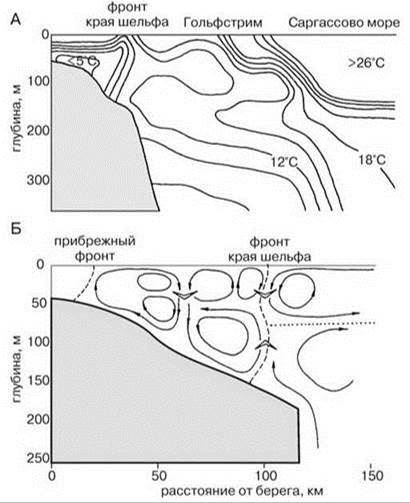
Рис. 5.27. Фронт края шельфа.
А — у атлантического побережья Северной Америки. По Longhurst (1998); Б — в западной части Берингова моря. По В.П. Шунтову (2001).
Непрерывное поступление биогенов в фотический слой поддерживает на протяжении всего вегетационного периода первичную продукцию в прибрежной водной массе на уровне, характерном для периода вспышки фитопланктона в более мористых районах. Это приводит к существенным различиям величин первичной продукции: в прибрежной зоне она во много раз выше. Лишь незначительную часть этой продукции потребляет растительноядный зоопланктон прибрежной водной массы. Так, в восточной части Берингова моря даже в период максимального развития он выедает менее 6% первичной продукции (Флинт, 2005). Часть этой продукции потребляет растительноядный зоопланктон неритической водной массы: в прилежащих к прибрежному фронту районах его биомасса на порядок больше, чем в удалении от фронта.
Прибрежной водной массой ограничено и массовое развитие донных макрофитов (фиталь). Какую-то часть (вероятно, существенную) продукции фитопланктона и бентосных растений потребляет верхнесублиторальный бентос, однако определить долю этого потребления в балансе всей экосистемы весьма сложно. Различия в функционировании экосистем по разные стороны прибрежного фронта приводят и к различиям их биот. С биогеографической точки зрения проекция этой границы на дно соответствует границе верхней и нижней сублиторали.
Фронт края шельфа. Изучение изображений, полученных со спутников, показало наличие квазипостоянных зон повышенной первичной продукции вдоль края шельфа, причём выраженность этих зон выше осенью. Наличие данных зон свидетельствует о существовании локальных апвеллингов (Рис. 5.27). Причины их возникновения ещё недостаточно ясны (существует несколько теорий). Наиболее аргументированным сейчас считают их возникновение в результате взаимодействия приливно-отливных и идущих вдоль склона течений с кромкой шельфа, особенно в местах со сложным рельефом. В результате образуются циклонические и антициклонические вихри в которых происходит подъём или опускание воды (Рис. 5.28). Апвеллинги кромки шельфа позволяют поддерживать уровень первичной продукции в районе фронта на уровне, характерном для периода вспышки фитопланктона в соседних районах. Эту продукцию потребляют концентрирующиеся здесь стада рыб, китов, ластоногих и даже птиц. Так, конюги-крошки (Aethia pusilla) питаются преимущественно (>85%) крупной растительноядной копеподой Neocalanus cristatus. Эта копепода наиболее массова в районе фронта, ширина которого обычно не превышает 20 км. Птицы дважды в сутки летают в район фронта от колонии на о. Сен-Поль (Рис. 5.29) (Флинт, 2005). Фронт края шельфа разделяет шельфовые (неритические) и океанические районы пелагиали. Проекция этого фронта на дно примерно соответствует границе сублиторали и батиали.
Расположение разных фронтов на примере Берингова моря показано на Рис. 5.29.
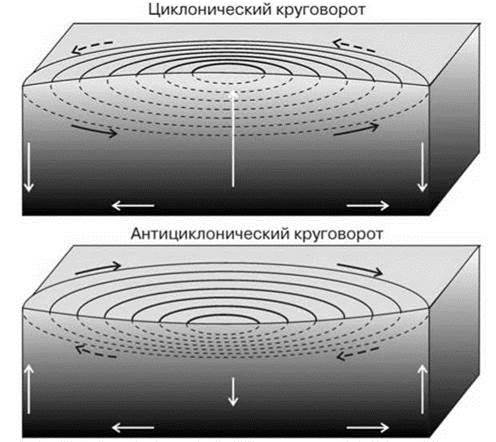
Рис. 5.28. Движение воды в циклонических и антициклонических круговоротах Северного полушария (в Южном полушарии движение обратное: в антициклонических круговоротах вода вращается против часовой стрелки, а в циклонических — по часовой стрелке).
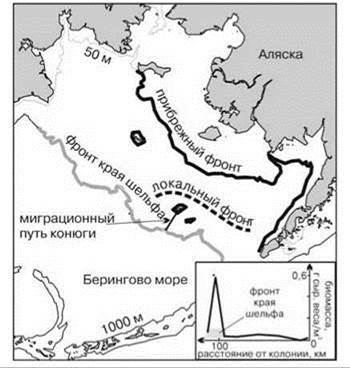
Рис. 5.29. Положение фронтов на восточном шельфе Берингова моря. По М.В. Флинту (2005).
На врезке — распределение Neocalanus cristatus вдоль миграционного пути конюги.
Океанические фронты (Рис. 5.30). Разделяют области распространения вод и водных масс. Ранг этих фронтов (а, следовательно, и границ) определяет размер, устойчивость и выраженность водных масс, которые они разделяют. Наиболее важны полярные фронты, расположенные в районах, где главный термоклин выходит на поверхность. Для них также характерны высокая продуктивность, благодаря интенсивно происходящему вертикальному перемешиванию, на малых глубинах охватывающему всю толщу воды от поверхности до дна.
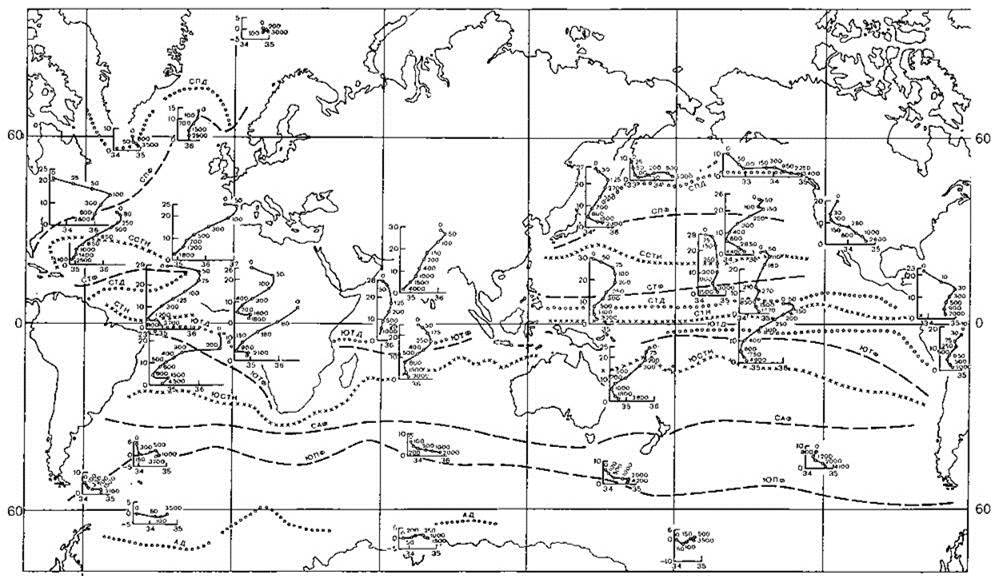
Рис. 5.30. Структура вод и разделяющие их фронты (пунктир) (по: В.А. Буркову, Р.П. Булатову, В.Г. Нейману (1973) из В.А. Буркова (1980).
АД — антарктическая дивергенция, САФ — субантарктический фронт (субтропическая конвергенция), СПД — субполярная дивергенция, СПФ — северный полярный фронт, ССТК — северная субтропическая конвергенция, СТД — северная тропическая дивергенция, СТК — северная тропическая конвергенция, СТФ — северный тропический фронт, ЮПФ — южный полярный фронт (антарктическая конвергенция), ЮСТК — южная субтропическая конвергенция, ЮТД — южная тропическая дивергенция, ЮТФ — южный тропический фронт. Вертикальная структура водных масс иллюстрирована TS-кривыми в точках, где помещено начало координат.